Bayağı tavşan
Bayağı tavşan[lower-alpha 1], yabani tavşan[lower-alpha 1] ya da sadece tavşan[lower-alpha 1] (Lepus europaeus), tavşangiller (Leporidae) familyasından kısa mesafede çok hızlı koşabilen bir tavşan türü. Avrupa ve Asya'nın bir böülümünde yaşar. En büyük tavşan türlerinden biridir ve ılıman açık araziye uyum sağlamıştır. Otçuldur ve otların yanı sıra özellikle kışları olmak üzere ağaç kabukları, taze dallar ve tomurcuklarla beslenir. Doğada büyük yırtıcı kuşlar, köpekgiller ve kedigiller tarafından avlanırlar. Uzun ve kuvvetleri bacakları ve büyük burun delikleri sayesinde avlanmaktan kaçmak için yüksek hızlı uzun süreli koşulara dayanırlar.
| Bayağı tavşan | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
-cropped.jpg.webp) | ||||||||||||||
| Korunma durumu | ||||||||||||||
| Bilimsel sınıflandırma | ||||||||||||||
| ||||||||||||||
(Pallas, 1778) | ||||||||||||||
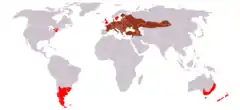 Yaşam alanları (koyu kırmızı:doğal, kırmızı:yeniden kazandırılan) | ||||||||||||||

Genellikle gececil olan ve doğası gereği ürkek olan tavşan ilkbaharda davranışını değiştirir ve gündüzleri açık alanlarda birbirlerini kovalarken görülebilir. Bu ilkbahar coşkunluğunda bazen birbirlerine patileri ile vururlar. Bu genellikle erkekler arasında bir rekabet değildir ancak bir dişinin erkeğe vurması ya onun henüz çiftleşmeye hazır olmadığını gösterir ya da erkeğin kararlılığını denemek için yapılır. Dişiler toprak içine kazılan bir oyuktan çok çukur alanlarda yuvalanırlar ve yavrular doğar doğmaz faaldirler. Dişiler bir kerede üç ila dört yavru doğurabilir ve yılda üç kere doğum yapabilirler. Tavşanlar on iki yıla kadar yaşayabilir. Üreme mevsimi Ocak ayından Ağustos ayına kadar sürer.
Bayağı tavşan yaygın dağılımı ve kısmen bol bulunması nedeniyle Dünya Doğa ve Doğal Kaynakları Koruma Birliği tarafından asgari endişe altındaki türler arasında listelenmiştir. Ancak kısmen tarım uygulamalarındaki değişiklik nedeniyle Avrupa anakarasında popülasyonu 1960'lardan beri azalma yönündedir. Yabani tavşan yüzyıllardır insanlar tarafından avlanmaktadır ve bir yılda vurulan tavşan sayısı beş milyonu geçmektedir. Tavşan bazı kültürlerde geleneksel olarak doğurganlık sembolü olarak görülmektedir.
Özellikleri
Bayağı tavşan Tavşangiller familyasının diğer üyeleri gibi hızlı koşan bir memelidir. Gözleri kafasını üstünde yukarıda konumlanmıştır; uzun kulakları ve esnek bir boynu vardır. Dişleri sürekli büyür. İlk kesici dişleri kemirmek için değişiklik göstermiş iken ikinci kesici dişleri ise kazık gibidir ve işlevsel değildir. Kesici dişler ile kaba bitkileri çiğnemek için uyum sağlamış azı dişleri arasında diastema yani boşluk vardır. Diş formülü 2/1, 0/0, 3/2, 3/3'tür.[3][4] Lepus cinsi tavşanların koyu uzuv kasları açık alanda hızlı ve sürekli koşmaya uyum sağlamıştır. Buna karşın pamuk kuyruk tavşanlar bitkilerin daha çok olduğu yerlerde kısa süreli hızlar için uyum sağlamışlardır.[5][6] Bayağı tavşanlarda hızlı koşmaya uyum sağlamanın diğer göstergeleri geniş burun delikleri ve daha büyük olan kalplaridir.[5] Avrupa ada tavşanı ile kıyaslandığında bayağı tavşanın orantısal olarak midesi ve çekumu daha küçüktür.[7]
Bayağı tavşan tavşanımsıların en büyüklerinden biridir. Kafası ve gövde uzunluğu 60 ila 75 cm. arasında değişir. Kuyruk uzunluğu da 7,2 ila 11 cm. arasındadır. Vücut ağırlığı genellikle 3 ila 5 kg. arasındadır.[8] Uzu kulakları 9,4 ila 11 cm. arasında uzunluğa sahiptir. Arka ayaklarının uzunluğu da 14 ila 16 cm. arasında değişir.[9] Kafatasındaki burun kemikleri kısa ama geniş ve kalındır. Supraorbital sırtın ön ve arka lobları iyi gelişmiştir ve lakrimal kemik güzyuvarının ön duvarından dışarıya doğru fırlar.[8]
Kürkü sırtında kırçıllı boz-sarı; omuzlarda, bacaklarda, ense ve boğazda kızıl kahverengi; alt kısmında beyaz ve kuyruk ile kulak uçlarında da siyahtır.[9] Sırttaki kürk genellikle vücudun diğer kısımlarında nazaran daha uzun ve daha kıvrıktır.[5] Aynı cinsteki diğer türlerin aksine bayağı tavşanın kürkü kış aylarında tamamen beyaza dönmez[9] ancak başın yanlarında ve kulakların tabanında beyaz bölgeler gelişir; kalçaa ile kıç bölgesi de biraz grileşebilir.[5]
Dağılımı ve yaşam alanı

Bayağı tavşan Kıta Avrupasının çoğunda ve Asya'nın bir bölümünde endemiktir. Dağılımı İspanya'nın kuzeyinden İskandinavya'nın güneyine, Doğu Avrupa'ya ve Güneybatı Asya'nın kuzey kısımları ile Orta Asya'ya kadar uzanır. Bu alan Sibirya'ya doğru genişlemektedir.[5] Britanya'ya yaklaşık 2000 yıl kadar önce Romalılar tarafından sokulmuş olmaları mümkündür çünkü daha önce tarihli yerleşimlerde haklarında bir kayıt yoktur. Lepus timidusun tek Lepus cinsi tavşan türü olduğu İrlanda'da yokturlar. Bazı Akdeniz adalarında da kayda düşmemiş sokulmalar olmuş olması muhtemeldir.[10] Kuzey Amerika (Ontario, New York, Pensilvanya'da başarısız olmuş, Massachusetts ve Connecticut), Güney Konisi (Brezilya, Arjantin, Uruguay, Paraguay), Bolivya, Şili, Peru, Falkland Adaları, Avustralya, Yeni Zelanda ve Rusya'nın güney Pasifik kıyılarına av hayvanı olarak sokulmuşlardır.[5][9][11]
Bayağı tavşanlar asıl olarak sığınak için kullandıkları kısmen çalılık ya da fundalıklı açık arazide yaşar. Çok uyumludurlar ve karma tarım alanlarında çok çabuk üreyebilirler.[5] Çekya'da yapılan bir araştırmaya göre 200 m. rakımın altında, yıllık 40 ila 60 gün kar kaplı olan yerlerde, yıllık yağış miktarının 400 ila 700 mm. olduğu yerlerde ve yıllık ortalama hava sıcaklığının 10 °C olduğu yerlerde bayağı tavşan popülasyonunun daha yüksek yoğunluğa sahip olduğunu göstermiştir. İklim açısından kışı hafif geçiren ılık ve yağışsız bölgelerde popülasyon yoğunluğunun daha yüksek olduğu da kaydedilmiştir.[12] Polonya'da bayağı tavşanların ormana kıyısı daha az bulunan yerlerde daha yoğun olduğu gözlemlenmiştir; bunun sebebi tilkilerin ormanı saklanmak için kullanmalarıdır. Açık alanda saklanabilecek yerlere ihtiyaç duyarlar çünkü bu alanlar gereksinim duydukları besinleri sağlar ve açık alanlarda yoğunlukları daha azdır. Toprağın yoğun olarak işlenmesi yavru tavşanların ölüm oranının artmasına neden olur.[13]
Büyük Britanya'da daha çok verimli tarlalarda, özellikle de nadasa bırakılan ya da buğday ve şeker pancarı ekili alanlarda görülürler. Özellikle otu bol meralarda, kısmen verimli tarım ürünleri ve kısmen de fundalıklar bulunan yerlerde sayıları artış gösterir. Tilkilerin veya şahinlerin çok bulunduğu yerlerde daha az sıklıkla görülürler. Her ne kadar Avrupa ada tavşanları ile çok az etkileşimleri olsa da ve aralarında saldırganlık davranışları görülmese de[14] ada tavşanı popülasyonlarının çok olduğu yerlerde de daha az sayıda oldukları görülür.[15] Bayağı tavşanlar sayıları çoğaldığında avlanılsa da az görüldükleri yerlerde pek avlanmazlar.[15]
Davranışı ve yaşam döngüsü
.jpg.webp)
Bayağı tavşanlar gececildir ve zamanlarının üçte birini yiyecek arayarak geçirirler.[5] Gündüzleri kısmen gizlenebildikleri yerdeki çöküntülerde geçirirler. Saatte 70 km. hızla koşabilirler ve karşılarına çıkan avcılardan daha hızlı koşarak kurtulmaya çalışırlar. Genel olarak asosyal oldukları düşünülse de küçük ya da büyük gruplar hâlinde de görülebilirler. Belirli bir bölgeyi sahiplenen hayvanlardan değildirler ve ortak yaşam alanları 300 hektar civarında bir bölgedir. Tavşanlar birbirleri arasında çeşitli görsel sinyallerle iletişim kurarlar. İlgilerini göstermek için kulaklarını dikerken diğerlerini uzak kalsınlar diye kulaklarını indirerek uyarırlar. Kendi türlerinden birine meydan okumak için ön ayaklarını kullanırlarken avcıları haber vermek için arka ayaklarını kullanırlar. Korktuğunda ya da bir yeri acıdığında tiz bir ses çıkarır; dişiler yavrularını çağırmak için gırtlaktan boğuk sesler çıkarır.[9] Bayağı tavşanlar on iki yıl kadar yaşarlar.[16]

Beslenme
Bayağı tavşanlar asıl olarak otçuldur. Yabani otlarla beslenirler ancak tarımın yaygınlaşması sonucu tercih ettikleri besinler olmadığında tarım mahsulleriyle de beslenmeye başlamışlardır.[16] İlkbahar ve yaz mevsimlerinde soya, yonca ve gelinciklerin[17] yanı sıra otlarla da beslenirler.[9] Sonbahar ve kış mevsimlerinde genel olarak buğday tercih etselerde ayrıca avcıların onların için bıraktıkları şeker pancarı ve havuçlarla da beslenirler.[17] Kışın aynı zamanda genç meyve ağaçlarının ve çalılıkların kabuklarını, sürgün ve tomurcuklarını da yerler.[9] Daha çekici besin kynakları varsa tahılları pek tercih etmezler; sadece lif yerine yüksek enerji veren besinleri tercih ederler.[18] Ağaç dalcıklarını ve sürgünleri yerken kabukları ayırıp çözünebilir karbonhidratları içeren damar dokulara ulaşırlar. Avrupa ada tavşanı ile kıyaslandığında sindirim oranlaarı benzer olmasına karşın bayağı tavşanda besin bağırsaklardan çok daha hızlı geçer.[7] Bazen sindirilmemiş proteinleri ve vitaminleri geri kazanmak için kendi yeşil dışkı parçalarını yedikleri görülür.[8] İki ila üç erişkin bayağı tavşan bir koyunun yediğinden daha fazla besin yiyebilir.[9]

Bayağı tavşanlar gruplar hâlinde beslenirler. Grup beslenmesinin yararı diğer tavşanların tetikte olduğunu bilerek beslenmek için daha çok zaman ayırabilmektir. Yine de besin kaynaklarının dağılımı bu yararları etkiler. Besin geniş bir alana yayıldıysa tüm tavşanlar ulaşabilir. Ancak besin dar bir alanda ise yalnızca baskın tavşanlar ulaşabilir. Küçük gruplarda baskın tavşanlar besinleri savunma yönünden daha başarılıdır ancak daha çok tavşan gruba katıldıkça zamanlarının çoğunu diğer tavşanları uzaklaştırmak için harcarlar. Grup ne kadar büyük olursa baskın tavşanların yeme için ayırdıkları süre azalır. Bu arada baskın tavşanlar meşgulken diğerleri besine ulaşabilir. Bu nedenle besin dar bir alanda ise gruptakilerin hepsi daha kötü beslenir.[19]


Üreme
Bayağı tavşanın üreme mevsimi uzundur ve Ocak ayından Ağustos ayına kadar sürer.[20][21] Dişiler üreme mevsimi boyunca gebe olarak görülebilir ve erkekler de Ekim ile Kasım ayları dışında çiftleşebilir. Bu ara dönemden sonra erkeklerin testisleri büyür ve faaliyeti artarak yeni üreme döngüsünün başını haber verir. Üreme organlarının tam işlevlerine kavuşması Aralık ile Şubat aylarında sürer. Ovülasyon başlamadan çiftleşme başlar ve yılın ilk gebelikleri genellikle tek fetus ile sonuçlanır ancak düşükler yaygın olarak görülür. Üreme eyleminin zirve yaptığı Mart ve Nisan aylarında tüm dişiler gebe kalabilir ve çoğunluğu üç ya da daha fazla fetuslü gebeliktir.[21]
Bayağı tavşanın çiftleşme sistemi çok eşliliktir.[22] Dişilerin üreme döngüsü altı haftadır ve her seferinde yalnızca birkaç saat çiftleşmeye açık olmaları erkek tavşanlar arasında büyük bir rekabete neden olur.[20] Üreme mevsiminin en yoğun olduğu dönemde gözlenen fenomene "Mart Çılgınlığı" adı verilir[21] ve bu dönemde normalde geceleri aktif olan erkekler gündüzleri de faal duruma geçerler. Baskın erkeklerin rakiplerine boyun eğdirmesinin yanı sıra dişiler de eğer çiftleşmeye hazır değilse peşinden gelen erkek tavşanları kovalar. Dövüşler şiddetli geçebilir ve kulaklarda sayısız yara izi bırakabilir.[20] Bu dövüşlerde tavşanlar iki ayaklarının üstünde dikilerek ön patileriyle birbirlerine saldırırlar; boks olarak bilinen bu davranışın sanıldığının aksine erkekler arasında değilde genellikle bir dişi ile erkek arasında olduğu anlaşılmıştır.[9][23] Dişi tavşan çiftleşmeye hazır olduğunda açık alanda koşmaya başlayarak peşine düşen erkeklerin dayanıklılığını ölçmeye yarayan bir testi başlatır. En dayanıklı erkek tavşan kaldığında dişi durarak onun çiftleşmeye başlamasına izin verir.[20] Dişiler Mayıs, Haziran ve Temmuz aylarında gebe kalabilmeye devam etseler de erkeklerde testosterone üretimi azalır ve cinsel davranışlar daha az belirgin hâle gelir. Bir batında doğan yavru sayısı üreme mevsimi kapanmaya yaklaştıkça azalır ve Ağustos ayından sonra gebelik olmaz. Erkeklerin testisleri küçülmeye başlar ve Eylül'de sperm üretimi durur.[21]

Dişiler yerdeki çukurlarda doğum yaparlar. Dişi tavşan 41 ila 42 gün süren gebelik sonucu yılda üç kere yavrulayabilir. Doğumda yavruların ortalama ağırlığı 130 g. civarındadır.[24] Yavrular tamamen kürklü olarak doğarlar ve doğar doğmaz yuvadan ayrılmaya hazırdırlar; bu yer altında korunaklı bir tavşan yuvasında olmamalarına karşın gelişmiş bir adaptasyondur.[9] Gündüzleri etrafa dağılan yavrular akşamları doğdukları yere yakın bir yerde toplanırlar. Anneleri günbatımından sonra onları emzirmek için yanlarına gelir; yavruların yaklaşık beş dakika boyunca süt emerler ve bu sırada idrarlarını yapar, annelerini de bu sıvıyı yalar. Daha sonra bir koku izi bırakmamak için zıplayarak uzaklaşır ve yavrular da tekrar dağılırlar.[9][25] Yavrular iki haftalıktan itibaren katı besinleri yemeye başlayabilir ve dört haftalıktan itibaren sütten kesilirler.[9] Hem dişi hem de erkek yavrular genel olarak çevrelerini keşfetseler de[26] biyolojik dağılım erkeklerde daha fazla olmaya meyillidir.[22][27] Dişiler cinsel olgunluğa yedi ila sekiz ayda erişirken erkeklerde bu süre altı aydır.[16]
Sağlık ve ölüm
.jpg.webp)
Bayağı tavşanlar büyük tavşanlardır ve erişkinleri yalnızca köpekgiller, kedigiller ve en büyük yırtıcı kuşlar tarafından avlanabilirler.[8] Polonya'da ilkbahar aylarında, daha küçük hayvanların az bulunduğu zamanlarda tilkilerin yediklerinin %50'sini tavşanların oluşturduğu görülmüştür; bayağı tavşanlarının ölüm oranlarının %50'si de tilkiler tarafından avlanmaları nedeniyledir.[28] İskandinavya'da kızıl tilki popülasyonunu azaltan bir uyuz salgını sonrasında bayağı tavşan sayılarının arttığı ve tilkilerin sayısının tekrar artmasıyla da tekrar eski sayılarına döndüğü gözlemlenmiştir.[29] Kaya kartalı Alplerde, Karpatlarda, Apenin Dağlarında ve İspanya'nın kuzeyinde bayağı tavşanları avlar.[30] Kuzey Amerika'da en çok tavşan avlayan avcılar tilkiler ve kır kurtlarıdır ancak ücra bölgelerde vaşakların da tavşan avladığı görülür.[24]
Bayağı tavşanlarda hem dış hem de iç parazitler bulunur. Bir araştırma Slovakya'da yaşayan bayağı tavşanların %54'ünde yuvarlak solucanlar, %90'ının üzerinde de koksidyumlar tespit edilmiştir.[31] Avustralya'da bayağı tavşanların dört yuvarlak solucan türü, altı koksidyum türü, çeşitli karaciğer kelebekleri ve iki şerit türü ile enfekte oldukları bildirilmiştir. Ayrıca tavşan piresi (Spilopsyllus cuniculi), tavuk piresi (Echidnophaga myrmecobii), bitler (Haemodipsus setoni ve H. lyriocephalus), ve maytlara (Leporacarus gibbus) konak oldukları da bildirilmiştir.[32]
Tavşan hemorajik hastalığına (RHD - "rabbit haemorrhagic disease") neden olana benzer bir calicivirüs tarafından neden olunan Avrupa boz tavşan sendromu (EBHS - "European brown hare syndrome") aynı şekilde ölümcüldür ama bu iki memeli türü arasında bulaşmaz.[33] Tavşanların karşılaştığı diğer tehditler ölümlerin ana kaynakları olan pastörelloz, yersinia psödotüberkülozu, koksidiyoz ve tularemidir.[34]
Ekim 2018'de miksomatoz virüsünün mutsyona uğramış bir formunun Büyük Britanya'da bayağı tavşanlara geçmiş olabileceği bildirilmiştir. Tavşanlarda nadir görülse de İspanya'da önemli sayıda tavşanın ölümüne neden olmuştur.[35][36]
Taksonomi ve genetik
.jpg.webp)
Bayağı tavşan ilk olarak 1778 yılında Alman zoolog Peter Simon Pallas tarafından tanımlanmıştır.[37] Lepus (Latince "tavşan"[38]) cinsinde diğer 31 tür ile birlikte sınıflandırılır.[39] Diğer tavşanımsılardan daha uzun bacakları, daha geniş burun delikleri ve yaşına göre erken gelişen yavrularıyla ayırt edilirler.[5] Lepus corsicanus, Lepus castroviejoi ve Lepus granatensis bir zamanlar bayağı tavşanın alt türleri olarak kabul edilmekteydi ancak DNA dizileme ve morfolojik analizler bunları ayrı bir tür olduğunu desteklemektedir.[40][41]
Lepus capensis ile bayağı tavşanın aynı tür olup olmadığı konusu tartışmalıdır. 2005 yılında yapılmış bir nükleer gen havuzu araştırması aynı tür olduklarını önerirken[42] 2006 yılında aynı hayvanlar üzerinde yapılan bir mitokondriyal DNA araştırmasında ise sonuç olarak bu hayvanların ayrı türler sayılacak kadar genetik olarak ayrıldıkları görüşü verilmiştir.[43] 2008 yılında yapılan bir araştırma ise Lepus cinsi söz konusu olduğunda ve hızlı evrimleşmeleri de göz önüne alındığında tür tanımı için yalnızca mtDNA sonuçlarına değil aynı zamanda nükleer gen havuzuna da bakmanın gerekli olduğunu iddia etmiştir. L. capensi ile L. europaeus arasındaki genetik farklar gerçek bir ayrışmadan çok coğrafi olarak ayrılmalarının sonucu olması muhtemeldir. Yakın Doğu'da tavşan popülasyonlarının birbirine karıştığı ve gen akışına maruz kaldıkları tahmininde bulunulmuştur.[44] 2008 yılında yapılan bir başka araştırma ise bir tür kompleksinin var olup olmadığına karar verebilmek için daha fazla araştırma yapılması gerektiğini önerir.[45] Bayağı tavşan daha fazla verinin bu önermeyle ters düşmesine kadar tek bir tür olarak sınıflandırılmaya devam etmektedir.[16]
Kladogenetik analiz bayağı tavşanın Pleistosen Çağı'nda son buzul döneminde Güney Avrupa (İtalya Yarımadası ve Balkanlar) ve Anadolu'daki sığınaklar yoluyla hayatta kaldığını göstermektedir. Orta Avrupa'da daha sonradan kolonileşmeleri insanlık kaynaklı çevresel değişiklikler nedeniyle olmuş görünmektedir.[46] Günümüz popülasyonlarında genetik çeşitlilik yüksektir ama inbred çiftleşmenin izleri görülmemektedir. Gen akışı erkeklere doğru temayüllü görünse de genelde popülasyonlar anasoylu yapıdadır. Almanya'nın Kuzey Ren-Vestfalya bölgesindeki tavşanlarda özellikle yüksek oranda genetik çeşitlilik olduğu görünmektedir. Yine de sınırlanmış gen akışının tecrit altında kalmış popülasyonlarda genetik çeşitliliği azaltabilmesi mümkündür.[47]
Tarihsel olarak 30 kadar bayağı tavşan alt türü tanımlanmıştır ancak statüleri tartışmalıdır.[5] Bu alt türler kürk rengi, vücut boyutları, dış gövde ölçüleri, kafatası morfolojisi ve diş şekillerindeki farklılıklar yoluyla ayırt edilirler.[48]
IUCN kırmızı kitabında Hoffmann ve Smith'e (2005) göre on altı alt tür listelenmiştir:[16]
- Lepus europaeus caspicus
- L. e. connori
- L. e. creticus
- L. e. cyprius
- L. e. cyrensis
- L. e. europaeus
- L. e. hybridus
- L. e. judeae
- L. e. karpathorum
- L. e. medius
- L. e. occidentalis
- L. e. parnassius
- L. e. ponticus
- L. e. rhodius
- L. e. syriacus
- L. e. transsylvanicus
Chapman ve Flux tarafından tavşanımsılar üzerine olan kitaplarında yukarıda belirtilen alt türlere ek olarak (L. e. connori, L. e. creticus, L. e. cyprius, L. e. judeae, L. e. rhodius ve L. e. syriacus haricinde) aşağıdakilerle birlikte "çok değişken statülü" yirmi dokuz alt tür listelenmiştir:[5]
- L. e. alba
- L. e. argenteogrisea
- L. e. biarmicus
- L. e. borealis
- L. e. caspicus
- L. e. caucasicus
- L. e. flavus
- L. e. gallaecius
- L. e. hispanicus
- L. e. hyemalis
- L. e. granatensis
- L. e. iturissius
- L. e. kalmykorum
- L. e. meridiei
- L. e. meridionalis
- L. e. niethammeri
- L. e. niger
- L. e. tesquorum
- L. e. tumak
İnsanlarla ilişkileri
Korunma durumu
Bayağı tavşan Avrupa ve Asya'nın batısında geniş bir alana dağılmıştır ve dünya üzerinde bazı ülkelere özellikle av hayvanı olarak sokulmuştur. Genel olarak doğal dağılımının olduğu bölgede kısmen yüksek popülasyona sahiptir[47] ancak 1960'lardan beri bazı bölgelerde popülasyonlarının azalmaya meyilli olduğu belirtilmiştir. Bunun nedeni olarak tarımsal uygulamaların yoğunlaşması gösterilmektedir.[49] Bayağı tavşan uyum sağlayabilen bir türdür ve yeni yaşam alanlarında geçebilir ancak daha çok bitki ve ot çeşitliliği olan yerlerde daha gelişebilir.[16] Bayağı tavşan bazı bölgelerde zararlı hayvan olarak kabul edilmektedir; kış aylarında alternatif besin kaynaklarının az olduğu yerlerde tarım mahsullerine ve genç ağaçlara daha çok zarar verebilmektedir.[9]
Dünya Doğa ve Doğal Kaynakları Koruma Birliği bayağı tavşanın korunma durumunu asgari endişe altında olarak belirlemiştir. Ancak düşük popülasyon yoğunluğu olan bölgelerde gen havuzunun azalması ile inbred çiftleşmenin daha mümkün olması nedeniyle yerel olarak soylarının tükenmesine hassas türler arasında olabilmektedirler. İspanya'nın kuzeyi ile Yunanistan'da bölge dışından getirilen tavşanların bölgesel gen havuzuna bir tehdit oluşturduğu görülmüştür. Buna karşılık olarak İspanya'da korunaklı bölgede yetiştirme programı başlatılmış ve bazı tavşanların bir bölgeden diğerine yerleştirilmesinin genetik çeşitliliği artırdığı görülmüştür.[16]
Bern Konvansiyonu bayağı tavşanı koruma altında türler arasında listeler.[15] Norveç, Almanya, Avusturya ve İsviçre gibi çeşitli ülkeler[16] bayağı tavşanı kendi "kırmızı liste"lerinde "neredeyse tehdit altında" ya da "tehdit altında" türler arasında gösterir.[50]
Notlar
- Türk Dil Kurumu Güncel Türkçe Sözlüğünde "tavşan" sözcüğünün tanımında Lepus europeus bilimsel adı verilmektedir. Bazı kaynaklarda "bayağı tavşan"[1] olarak geçerken başka kaynaklarda "yabanitavşan"[2] olarak kullanılmaktadır.
Kaynakça
- "tavşan". AnaBritannica. 20. Ana Yayıncılık. 1990. s. 453.
- Demirsoy, Ali (1992). Yaşamın Temel Kuralları Omurgalılar/Amniyota (Sürüngenler, Kuşlar ve Memeliler) Cilt - III / Kısım - II. Ankara: Meteksan. s. 695. ISBN 975-7746-02-9.
- Alves, Paulo C.; Ferrand, Nuno; Hackländer, Klaus (2007). Lagomorph Biology: Evolution, Ecology, and Conservation. Springer Science & Business Media. s. 1. ISBN 978-3-540-72446-9.
- Leach, Michael (2008). Hare. The Rosen Publishing Group. s. 4. ISBN 978-1-4358-4997-6.
- Chapman, Joseph A.; Flux, John E. C. (1990). Rabbits, Hares and Pikas: Status Survey and Conservation Action Plan. IUCN. ss. 62, 76-78. ISBN 978-2-8317-0019-9. 25 Aralık 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Şubat 2020.
- Schnurr, D. L.; Thomas, V. G. (1984). "Histochemical properties of locomotory muscles of European hares and cottontail rabbits". Canadian Journal of Zoology. 62 (11). ss. 2157-2163. doi:10.1139/z84-313.
- Stott, P. (2008). "Comparisons of digestive function between the European hare (Lepus europaeus) and the European rabbit (Oryctolagus cuniculus): Mastication, gut passage, and digestibility". Mammalian Biology – Zeitschrift für Säugetierkunde. 73 (4). ss. 276-286. doi:10.1016/j.mambio.2007.07.002.
- "Lepus europaeus European hare". Animal Diversity. University of Michigan. 14 Temmuz 2014 tarihinde kaynağından arşivlendi. Erişim tarihi: 11 Mayıs 2011.
- Naughton, D. (2012). The Natural History of Canadian Mammals. University of Toronto Press. ss. 235-238. ISBN 978-1-4426-4483-0.
- Corbet, G. B. (1986). "Relationships and origins of the European lagomorphs". Mammal Review. 16 (3–4). ss. 105-110. doi:10.1111/j.1365-2907.1986.tb00029.x.
- Bonino, N. A.; Cossios, D.; Menegheti, J. (2010). "Dispersal of the European hare, Lepus europaeus in South America" (PDF). Folia Zoologica Praha. 59 (1). ss. 9-15. doi:10.25225/fozo.v59.i1.a3.2010. 17 Eylül 2016 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 16 Eylül 2016.
- Pikula, J.; Beklová, M.; Holešovská, Z.; Treml, F. (2004). "Ecology of European brown hare and distribution of natural foci of Tularaemia in the Czech Republic". Acta Veterinaria Brno. 73 (2). ss. 267-273. doi:10.2754/avb200473020267.
- Panek, Manek; Kamieniarz, Robert (1999). "Relationships between density of brown hare Lepus europaeus and landscape structure in Poland in the years 1981–1995". Acta Theriologica. 44 (1). ss. 67-75. doi:10.4098/at.arch.99-7. ISSN 0001-7051.
- Alves, Paulo C.; Ferrand, Nuno; Hackländer, Klaus (2007). Lagomorph Biology: Evolution, Ecology, and Conservation. Springer Science & Business Media. s. 247. ISBN 978-3-540-72446-9.
- Vaughan, N.; Lucas, E.; Harris, S.; White, P. C. L. (2003). "Habitat associations of European hares Lepus europaeus in England and Wales: Implications for farmland management". Journal of Applied Ecology. 40 (1). ss. 163-175. doi:10.1046/j.1365-2664.2003.00784.x. JSTOR 827268.
- Hacklander, K.; Schai-Braun, S. (2019). "Lepus europaeus". The IUCN Red List of Threatened Species. s. e.T41280A45187424. doi:10.2305/IUCN.UK.2019-1.RLTS.T41280A45187424.en. 18 Temmuz 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Şubat 2020.
- Reichlin, T.; Klansek, E.; Hackländer, K. (2006). "Diet selection by hares (Lepus europaeus) in arable land and its implications for habitat management". European Journal of Wildlife Research. 52 (2). ss. 109-118. doi:10.1007/s10344-005-0013-3.
- Schai-Braun, S.C.; Reichlin, T. S.; Ruf, T.; Klansek, E.; Tataruch, F.; Arnold, W.; Hackländer, K. (2015). "The European hare (Lepus europaeus): A picky herbivore searching for plant parts rich in fat". PLoS ONE. 10 (7). s. e0134278. Bibcode:2015PLoSO..1034278S. doi:10.1371/journal.pone.0134278. PMC 4521881 $2. PMID 26230115.
- Monaghan, P.; Metcalfe, N. B. (1985). "Group foraging in wild brown hares: Effects of resource distribution and social status". Animal Behaviour. 33 (3). ss. 993-999. doi:10.1016/S0003-3472(85)80033-6.
- Holly, T. (2001). "Mad World of the European Hare". MacDonald, D. (Ed.). The New Encyclopedia of Mammals. Oxford: Oxford University Press. s. 710–711. ISBN 978-0-19-850823-6.
- Lincoln, G. (1974). "Reproduction and March madness in the brown hare, Lepus europaeus". Journal of Zoology. 174 (1). ss. 1-14. doi:10.1111/j.1469-7998.1974.tb03140.x. PMID 4468894.
- Bray, Y.; Devillard, S.; Marboutin, E.; Mauvy, B.; Péroux, R. (2007). "Natal dispersal of European hare in France" (PDF). Journal of Zoology. 273 (4). ss. 426-434. doi:10.1111/j.1469-7998.2007.00348.x. 6 Ekim 2016 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 28 Şubat 2020.
- Holley, A.J.F.; Greenwood, P.J. (1984). "The myth of the mad March hare". Nature. 309 (5968). ss. 549-550. Bibcode:1984Natur.309..549H. doi:10.1038/309549a0. PMID 6539424.
- Kurta, Allen (1995). Mammals of the Great Lakes Region. University of Michigan Press. s. 104. ISBN 978-0-472-06497-7.
- Broekhuizen, S.; Maaskamp, F. (1980). "Behaviour of does and leverets of the European hare (Lepus europaeus) while nursing". Journal of Zoology. 191 (4). ss. 487-501. doi:10.1111/j.1469-7998.1980.tb01480.x.
- Avril, A.; Letty, J.; Léonard, Y.; Pontier, D. (2014). "Exploration forays in juvenile European hares (Lepus europaeus): dispersal preludes or hunting-induced troubles?". BMC Ecology. Cilt 14. s. 16. doi:10.1186/1472-6785-14-6. PMC 3943402 $2. PMID 24568541.
- Avril, A.; Letty, J.; Léonard, Y.; Pérouxb, R.; Guitton, J.-S.; Pontier, D. (2011). "Natal dispersal of European hare in a high-density population". Mammalian Biology – Zeitschrift für Säugetierkunde. 76 (2). ss. 148-156. doi:10.1016/j.mambio.2010.07.001.
- Goszczyński, J.; Wasilewski, M. (1992). "Predation of foxes on a hare population in central Poland". Acta Theriologica. 37 (4). ss. 329-338. doi:10.4098/at.arch.92-33. ISSN 0001-7051.
- Lindstrom, Eric R.; Andren, Henrik; Angelstam, Per; Cederlund, Goran; Hornfeldt, Birger; Jaderberg, Lars; Lemnell, Per-Arne; Martinsson, Berit; Skold, Kent; Swenson, John E. (1992). "Disease reveals the predator: sarcoptic mange, red fox predation and prey populations". Ecology. 75 (4). ss. 1042-1049. doi:10.2307/1939428. JSTOR 1939428.
- Watson, Jeff; Brockie, Keith (1997). The Golden Eagle. A&C Black. s. 50. ISBN 978-0-85661-099-8. 19 Ocak 2017 tarihinde kaynağından arşivlendi. Erişim tarihi: 28 Şubat 2020.
- Dubinský, P.; Vasilková, Z.; Hurníková, Z.; Miterpáková, M.; Slamečka, J.; Jurčík, R. (2010). "Parasitic infections of the European brown hare (Lepus europaeus Pallas, 1778) in south-western Slovakia". Helminthologia. 47 (4). ss. 219-225. doi:10.2478/s11687-010-0034-7.
- "European hare". Agriculture: Pest animals. Agriculture Victoria. 2 Ağustos 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 7 Eylül 2016.
- Alves, Paulo C.; Ferrand, Nuno; Hackländer, Klaus (2007). Lagomorph Biology: Evolution, Ecology, and Conservation. Springer Science & Business Media. s. 263. ISBN 978-3-540-72446-9.
- Lamarque, F.; Barrat, J.; Moutou, F. (1996). "Principal diagnoses for determining causes of mortality in the European brown hare (Lepus europaeus) found dead in France between 1986 and 1994". Gibier Faune Sauvage. 13 (1). ss. 53-72.
- Yohannes, Lowe (11 Ekim 2018). "Hares could be wiped out, experts warn, as mystery deaths spark fears RHD-2 has 'jumped' from rabbits". The Telegraph. 15 Ağustos 2019 tarihinde kaynağından arşivlendi. Erişim tarihi: 28 Şubat 2020.
- "Concern over hare deaths". New Scientist. 20 Ekim 2018. s. 4.
- Pallas, Peter Simon (1778). Novae Species Quadrupedum e Glirium Ordine (Latince). Erlanger: Wolfgangi Waltheri. s. 30. 24 Şubat 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 24 Şubat 2020.
- "Lepus". Online Etymology Dictionary. 6 Ekim 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 15 Eylül 2016.
- Hoffman, R.S.; Smith, A.T. (2005). "Genus Lepus". Wilson, D.E.; Reeder, D.M (Edl.). Mammal Species of the World: A Taxonomic and Geographic Reference (3rd bas.). Johns Hopkins University Press. ss. 195-205. ISBN 978-0-8018-8221-0. OCLC 62265494. 8 Mayıs 2014 tarihinde kaynağından arşivlendi. Erişim tarihi: 4 Ekim 2020.
- Palacios, F. (1989). "Biometric and morphologic features of the species of the genus Lepus in Spain". Mammalia. 53 (2). ss. 227-264. doi:10.1515/mamm.1989.53.2.227.
- Riga, F.; Trocchi, V.; Randi, E.; Toso, S. (2001). "Morphometric differentiation between the Italian hare (Lepus corsicanus De Winton, 1898) and the European brown hare (Lepus europaeus Pallas, 1778)". Journal of Zoology. 253 (2). ss. 241-252. doi:10.1017/S0952836901000218.
- Ben Slimen, H.; Suchentrunk, F.; Memmi, A.; Ben Ammar Elgaaied, A. (2005). "Biochemical genetic relationships among Tunisian hares (Lepus sp.), South African Cape hares (L. capensis), and European brown hares (L. europaeus)". Biochemical Genetics. 43 (11–12). ss. 577-596. doi:10.1007/s10528-005-9115-6. PMID 16382363.
- Ben Slimen, H.; Suchentrunk, F.; Memmi, A.; Sert, H.; Kryger, U.; Alves, P. C.; Elgaaied, A. B. A. (2006). "Evolutionary relationships among hares from North Africa (Lepus sp. or Lepus spp.), Cape Hares (L. capensis) from South Africa, and brown hares (L. europaeus), as inferred from mtDNA PCR-RFLP and allozyme data". Journal of Zoological Systematics and Evolutionary Research. 44 (1). ss. 88-99. doi:10.1111/j.1439-0469.2005.00345.x.
- Ben Slimen, H.; Suchentrunk, F.; Ben Ammar Elgaaied, A. (2008). "On shortcomings of using mtDNA sequence divergence for the systematics of hares (genus Lepus): An example from Cape hares". Mammalian Biology. 73 (1). ss. 25-32. doi:10.1016/j.mambio.2007.02.003.
- Ben Slimen, H.; Suchentrunk, F.; Stamatis, C.; Mamuris, Z.; Sert, H.; Alves, P. C.; Kryger, U.; Shahin, A. B.; Ben Ammar Elgaaied, A. (2008). "Population genetics of Cape and brown hares (Lepus capensis and L. europaeus): A test of Petter's hypothesis of conspecificity". Biochemical Systematics and Ecology. 36 (1). ss. 22-39. doi:10.1016/j.bse.2007.06.014.
- Fickel, J.; Hauffe, H. C.; Pecchioli, E.; Soriguer, R.; Vapa, L.; Pitra, C. (2008). "Cladogenesis of the European brown hare (Lepus europaeus Pallas, 1778)" (PDF). European Journal of Wildlife Research. 54 (3). ss. 495-510. doi:10.1007/s10344-008-0175-x. hdl:10261/62899. 23 Temmuz 2018 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 24 Şubat 2020.
- Fickel, J.; Schmidt, A.; Putze, M.; Spittler, H.; Ludwig, A.; Streich, W. J.; Pitra, C. (2005). "Genetic structure of populations of European brown hare: Implications for management". Journal of Wildlife Management. 69 (2). ss. 760-770. doi:10.2193/0022-541X(2005)069[0760:GSOPOE]2.0.CO;2.
- Suchentrunk, F.; Mamuris, Z.; Sfougaris, A. I.; Stamatis, C. (2003). "Biochemical genetic variability in brown hares (Lepus europaeus) from Greece". Biochemical Genetics. 41 (5–6). ss. 127-140. doi:10.1023/A:1023354709392. PMID 12834043.
- Smith, R. K.; Jennings, N. V.; Harris, S. (2005). "A quantitative analysis of the abundance and demography of European hares Lepus europaeus in relation to habitat type, intensity of agriculture and climate". Mammal Review. 35 (1). ss. 1-24. doi:10.1111/j.1365-2907.2005.00057.x.
- Reichlin, T.; Klansek, E.; Hackländer, K. (2006). "Diet selection by hares (Lepus europaeus) in arable land and its implications for habitat management". European Journal of Wildlife Research. 52 (2). ss. 109-118. doi:10.1007/s10344-005-0013-3.
Ada tavşanı • Amami tavşanı • Ankara tavşanı • Avrupa ada tavşanı • Bayağı tavşan • Belçika tavşanı • Beveren tavşanı • Havana tavşanı • Hollanda cüce tavşanı • Hollanda lop tavşanı • Kaliforniya tavşanı • Pigme tavşan • Sumatra tavşanı • Viyana tavşanı • Yeni Zelanda beyaz tavşanı • Yeni Zelanda kırmızı tavşanı • İngiliz tavşanı | |||