Enzim katalizi
Enzim katalizi, biyolojik bir molekül olan "enzim" tarafından sağlanan, bir sürecin hızındaki artıştır. Enzimlerin çoğu proteindir ve bu tür işlemlerin çoğu kimyasal reaksiyonlardır. Enzim içinde, genellikle kataliz aktif bölge olarak adlandırılan lokalize bir bölgede gerçekleşir.
Çoğu enzim, ağırlıklı olarak proteinlerden, tek bir protein zinciri veya çok alt birimli bir kompleks içindeki bu tür birçok zincirden yapılır. Enzimler sıklıkla ayrıca metal iyonları veya kofaktör olarak bilinen özel organik moleküller (Ör. Adenosin trifosfat) gibi protein olmayan bileşenleri de içerir. Kofaktörlerin birçoğu vitaminlerdir ve vitaminler olarak rolleri, metabolizma içindeki biyolojik sürecin katalizinde kullanımlarıyla doğrudan bağlantılıdır. Biyokimyasal reaksiyonların katalizi, hücre için hayati önem taşımaktadır, çünkü bazı reaksiyonlar katalize edilmediğinde aşırı yavaş gerçekleşir. Protein evriminin bir itici gücü, bu tür katalitik aktivitelerin optimizasyonudur, ancak yalnızca en önemli enzimler, katalitik verimlilik sınırlarına yakın çalışır ve birçok enzim optimal değildir. Enzim katalizindeki önemli faktörler arasında genel asit ve baz katalizi, orbital yönlendirme, entropik kısıtlama, oryantasyon etkileri (yani kilit ve anahtar kataliz) ve ayrıca protein dinamiklerini içeren hareket etkileri bulunur.[1]
Enzim kataliz mekanizmaları değişiklik gösterse de prensip olarak diğer kimyasal kataliz türlerine benzer, önemli olan faktör reaktanları ürünlerden ayırmada enerji bariyer(ler)inin azalmasıdır[2]. Aktivasyon enerjisinin indirgenmesi, bu engeli aşan ve ürün oluşturabilen reaktif moleküllerin oranını arttırır. Sadece ürünler ve reaktanlar arasındaki enerji bariyerlerini azalttıkları için, enzimlerin her zaman her iki yönde reaksiyonları katalize etmeleri, bir reaksiyonu ileri sürdürememeleri veya denge pozisyonunu etkileyememeleri önemli bir ilkedir. Diğer katalizörlerde olduğu gibi, enzim de reaksiyon sırasında tüketilmez veya değiştirilmez, ancak tek bir enzimin birçok kataliz türü gerçekleştireceği şekilde geri dönüştürülür.
İndüklenmiş uyum
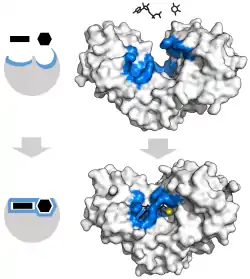
Enzim-substrat etkileşimi için klasik model, indüklenmiş uyum modelidir.[3] Bu model, enzim ve substrat arasındaki ilk etkileşimin nispeten zayıf olduğunu, ancak bu zayıf etkileşimlerin hızla enzimde bağlanmayı güçlendiren konformasyonel değişiklikleri indüklediğini ileri sürer.
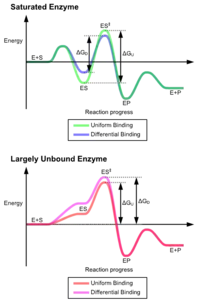
İndüklenmiş uyum mekanizmasının avantajları, güçlü enzim bağlanmasının stabilize edici etkisinden kaynaklanmaktadır. İki farklı substrat bağlama mekanizması vardır: güçlü substrat bağlamasına sahip tek tip ciltleme ve güçlü geçiş durumu bağlamasına sahip diferansiyel bağlama. Tek tip bağlanmanın stabilize edici etkisi, hem substratı hem de geçiş durumu bağlanma afinitesini arttırırken, diferansiyel bağlama yalnızca geçiş durumu bağlanma afinitesini artırır. Her ikisi de enzimler tarafından kullanılır ve reaksiyonun aktivasyon enerjisini en aza indirmek için evrimsel olarak seçilmiştir. Doymuş, yani yüksek afiniteli bir substrat bağlanmasına sahip enzimler, aktivasyon enerjisini azaltmak için farklı bağlanma gerektirirken, küçük substrat bağlı olmayan enzimler, diferansiyel veya tek tip bağlanma kullanabilir.[4]
Bu etkiler, çoğu proteinin, aktivasyon enerjisini azaltmak için diferansiyel bağlanma mekanizmasını kullanmasına yol açmıştır, bu nedenle çoğu substrat, geçiş durumundayken enzime yüksek afiniteye sahiptir. Diferansiyel bağlanma, indüklenmiş uyum mekanizması ile gerçekleştirilir - substrat önce zayıf bir şekilde bağlanır, ardından enzim, geçiş durumuna afiniteyi artırarak ve stabilize ederek konformasyonu değiştirir, böylece ona ulaşmak için aktivasyon enerjisini azaltır.
Bununla birlikte, uyarılmış uyum kavramının katalizi rasyonelleştirmek için kullanılamayacağını açıklığa kavuşturmak önemlidir. Kimyasal kataliz Ea'nin suda katalize edilmemiş reaksiyonuna göre azalması olarak tanımlanır. İndüklenmiş uyum, sadece enzimin kapalı formunda bariyerin daha düşük olduğunu gösterir, ancak bariyer azalmasının nedenini söylemez.
İndüklenmiş uyum, konformasyonel düzeltme okuma mekanizması yoluyla rekabet ve gürültünün varlığında moleküler tanımanın doğruluğu için faydalı olabilir.[5]
Alternatif bir reaksiyon yolunun mekanizmaları
Bu konformasyonel değişiklikler ayrıca, reaksiyonda değişecek olan substrattaki kimyasal bağlara yakın aktif bölgede katalitik kalıntılar oluşturur. Bağlanma gerçekleştikten sonra, bir veya daha fazla kataliz mekanizması, reaksiyon için alternatif bir kimyasal yol sağlayarak reaksiyonun geçiş durumunun enerjisini düşürür. "Bariyer üzerinden" katalizinin altı olası mekanizması ve "bariyer üzerinden" bir mekanizma vardır:
Yakınlık ve yönelim
Enzim-substrat etkileşimleri, reaktif kimyasal grupları hizalar ve bunları optimum bir geometride birbirine yakın tutar, bu da reaksiyonun hızını artırır. Bu, reaktanların entropisini azaltır ve bu nedenle, iki reaktant tek bir ürün haline geldiğinde toplam entropide bir azalma olduğundan, ekleme veya transfer reaksiyonlarını daha az elverişsiz hale getirir. Bununla birlikte, bu genel bir etkidir ve reaktiflerin "etkili konsantrasyonundaki" bir artıştan dolayı meydana geldiği ilavesiz veya transfer reaksiyonlarında görülür. Konsantrasyondaki artışların reaksiyon hızında nasıl artışlara yol açtığı düşünüldüğünde, esasen reaktanların daha konsantre olduklarında daha sık çarpıştıkları ve bu nedenle daha sık reaksiyona girdikleri anlaşılır. Enzim katalizinde, reaktiflerin enzime bağlanması, reaktanların konformasyonel alanını sınırlandırır, onları 'uygun oryantasyonda' ve birbirlerine yakın tutar, böylece çarpışmayı kolaylaştırmak için daha sık ve doğru geometri ile istenen reaksiyonu sağlar. "Etkili konsantrasyon", aynı çarpışma frekansını deneyimlemek için reaktantın çözelti içinde serbest olması gereken konsantrasyondur. Çoğu zaman, bu tür teorik konsantrasyonlar fiziksel olarak gerçekçi değildir ve gerçekleştirilmesi imkansızdır - bu, pek çok enzimin katalitik gücünün katalizlenmemiş duruma göre muazzam hız artışı sağladığının bir kanıtıdır.
| Örneğin: |
| Reaksiyon molekül içi ise benzer reaksiyonlar çok daha hızlı gerçekleşecektir. |
 |
| İçinde etkili yoğunlukta asetat intramoleküler reaksiyonda k 2 / k, 1 = 2 x 10 5 Molar olarak tahmin edilebilir. |
Bununla birlikte, modern hesaplama çalışmaları geleneksel yakınlık etkilerinin örneklerinin doğrudan enzim entropik etkileriyle ilişkilendirilemeyeceğini belirlediğinden, durum daha karmaşık olabilir.[6][7][8] Ayrıca, orijinal entropik önerinin[9], oryantasyon entropisinin katalize katkısını büyük ölçüde abarttığı bulunmuştur.[10]
Proton bağışçıları veya alıcıları
Proton vericileri ve alıcıları, yani asitler ve bazlar, geçiş durumunda gelişen yükleri stabilize etmek için protonları bağışlayabilir ve kabul edebilir. Bu, genel olarak geçiş durumları yüksek enerji durumları olduğundan ve bunları stabilize ederek bu yüksek enerji azaltılarak bariyeri düşürdüğünden, genel kataliz ilkesi olan enerji engellerini azaltma ilkesiyle ilgilidir. Enzim katalizinin biyolojik olmayan birçok katalize göre temel bir özelliği, hem asit hem de baz katalizinin aynı reaksiyonda birleştirilebilmesidir. Birçok abiyotik sistemde, asitler (büyük [H +]) veya bazlar (büyük konsantrasyonlu H + yutakları veya elektron çiftleri olan türler) reaksiyon hızını artırabilir; ama tabii ki ortam yalnızca bir toplam pH'a sahip olabilir (asitlik veya bazlık ölçüsü, alkalinite). Bununla birlikte, enzimler büyük moleküller olduğundan, hem asit gruplarını hem de bazik grupları aktif bölgelerinde substratlarıyla etkileşime girecek şekilde konumlandırabilirler ve her iki modu da yığın pH'ından bağımsız olarak kullanırlar.
Nükleofil ve/veya elektrofil gruplarını etkinleştirmek veya ayrılan grupları stabilize etmek için genellikle genel asit veya baz katalizi kullanılır. Glutamik ve aspartik asit, histidin, sistin, tirosin, lisin ve arginin ile serin ve treonin gibi asidik veya bazik gruplara sahip birçok amino asit bu aktif bölgede kullanılır. Ek olarak, karbonil ve amid N grupları içeren peptit omurgası sıklıkla kullanılır. Sistin ve Histidin, her ikisi de nötr pH'a yakın bir pKa'ya sahip oldukları ve bu nedenle protonları hem kabul edip hem de bağışlayabildikleri için çok yaygın olarak yer alırlar.
Asit / baz katalizini içeren birçok reaksiyon mekanizması, büyük ölçüde değiştirilmiş bir pKa olduğunu varsayar. Bu pKa değişikliği, kalıntının yerel ortamı aracılığıyla mümkündür.
| Koşullar | Asitler | Bazlar |
|---|---|---|
| Hidrofobik ortam | PKa'yı artırır | PKa'yı azaltır |
| Benzer yükün bitişik kalıntıları | PKa'yı artırır | PKa'yı azaltır |
| Tuz köprüsü (ve hidrojen bağ) oluşumu |
PKa'yı azaltır | PKa'yı artırır |
pKa, çözelti içinde bazik olan kalıntıların proton donörleri olarak hareket edebildiği ölçüde çevreleyen ortamdan da önemli ölçüde etkilenebilir ve bunun tersi de geçerlidir.
| Örneğin: |
| Serin proteazın katalitik üçlüsü |
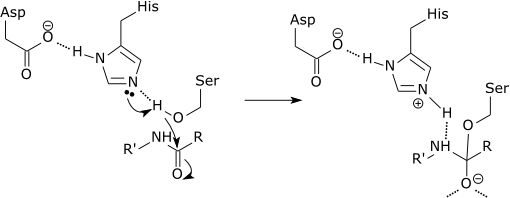 |
| Serin proteaz katalitik mekanizmasının ilk adımı, aktif bölgenin histidininin serin kalıntısından bir protonu kabul etmesini içerir. Bu, serini, substratın amid bağına saldırmak için bir nükleofil olarak hazırlar. Bu mekanizma, serinden (bir baz, pKa 14) histidine (bir asit, pKa 6), bazların yerel ortamı nedeniyle mümkün kılınan bir protonun bağışlanmasını içerir. |
PKa'ların modifikasyonunun elektrostatik mekanizmanın saf bir parçası olduğunu açıklığa kavuşturmak önemlidir.[11] Ayrıca, yukarıdaki örneğin katalitik etkisi esas olarak oksianyonun pKa'sının azalması ve histidinin pKa'sındaki artışla ilişkilidir; serinden histidine proton transferi önemli ölçüde katalize edilmez, çünkü bariyeri belirleyen oran değildir.[12]
Elektrostatik kataliz
Yüklü geçiş durumlarının stabilizasyonu, aktif bölgedeki ara ürünle iyonik bağlar (veya kısmi iyonik yük etkileşimleri) oluşturan kalıntılar yoluyla da olabilir. Bu bağlar, lisin, arginin, aspartik asit veya glutamik asit gibi amino asitlerde bulunan asidik veya bazik yan zincirlerden veya çinko gibi metal kofaktörlerden gelebilir. Metal iyonları özellikle etkilidir ve suyun pKa'sını onu etkili bir nükleofil yapacak kadar azaltabilir.
Sistematik bilgisayar simülasyon çalışmaları, elektrostatik etkilerin katalize en büyük katkıyı sağladığını tespit etti.[11] Bu 107 faktörü ile reaksiyonun hızını artırabilir.[13] Özellikle enzimin sudan daha polar bir ortam sağladığı ve iyonik geçiş durumlarının sabit dipollerle stabilize edildiği bulunmuştur. Bu, su moleküllerinin "yeniden düzenleme enerjisi" kullanması gereken sudaki geçiş durumu stabilizasyonundan çok farklıdır.[14] Bu nedenle kataliz, enzim polar gruplarının önceden organize edilmiş olması gerçeğiyle ilişkilidir.[15]
Bir enzimin aktif bölgesi tarafından uygulanan elektrostatik alanın büyüklüğünün, enzimin katalitik hız artışı ile oldukça ilişkili olduğu gösterilmiştir.[16][17]
Substratın bağlanması genellikle aktif bölgedeki suyu dışarıda bırakır, böylece yerel dielektrik sabitini bir organik solventinkine düşürür. Bu, yüklü / polar substratlar ile aktif bölgeler arasındaki elektrostatik etkileşimleri güçlendirir. Ek olarak, çalışmalar, aktif bölgeler hakkındaki yük dağılımlarının, katalize reaksiyonların geçiş durumlarını stabilize edecek şekilde düzenlendiğini göstermiştir. Birkaç enzimde, bu yük dağılımları görünüşte polar substratları bağlanma bölgelerine doğru yönlendirmeye hizmet eder, böylece bu enzimatik reaksiyonların oranları, görünür difüzyon kontrollü sınırlarından daha büyüktür.
| Örneğin: |
| Karboksipeptidaz katalitik mekanizması |
 |
| Dört yüzlü ara ürün, Zn2 + iyonu ile oksijen üzerindeki negatif yük arasında kısmi bir iyonik bağ ile stabilize edilir. |
Kovalent kataliz
Kovalent kataliz, substratın enzim aktif sahasındaki tortularla veya bir kofaktörle geçici bir kovalent bağ oluşturmasını içerir. Bu, reaksiyona ilave bir kovalent ara ürün ekler ve reaksiyonun sonraki geçiş durumlarının enerjisini azaltmaya yardımcı olur. Kovalent bağ, reaksiyonun sonraki bir aşamasında, enzimi yeniden oluşturmak için kırılmalıdır. Bu mekanizma, bir asil enzim ara maddesinin oluştuğu kimotripsin ve tripsin gibi proteazlar gibi enzimlerin katalitik üçlüsü tarafından kullanılır. Alternatif bir mekanizma, glikoliz sırasında aldolaz enziminde görüldüğü gibi, bir lizin kalıntısından serbest amin kullanılarak schiff bazı oluşumudur.
Bazı enzimler, reaktan molekülleri ile kovalent ara ürünler oluşturmak için piridoksal fosfat (PLP) veya tiamin pirofosfat (TPP) gibi amino asit olmayan kofaktörler kullanır.[18][19] Bu tür kovalent ara maddeler, aktif alan amino asit kalıntıları ile oluşturulan kovalent ara maddelerin stabilizasyona izin vermesine benzer şekilde, sonraki geçiş durumlarının enerjisini azaltma işlevi görür, ancak kofaktörlerin yetenekleri, enzimlerin amino asit yan kalıntılarının tek başına yapamayacağı reaksiyonları gerçekleştirmesine izin verir. Bu tür kofaktörleri kullanan enzimler, PLP'ye bağımlı enzim aspartat transaminazı ve TPP'ye bağımlı enzim piruvat dehidrojenazı içerir.[20][21]
Bir reaksiyon yolu için aktivasyon enerjisini düşürmek yerine, kovalent kataliz reaksiyon için alternatif bir yol sağlar (kovalent ara ürün yoluyla) ve bu nedenle gerçek katalizden farklıdır.[11] Örneğin, kimotripsin içindeki serin molekülüne kovalent bağın enerjisi, katalize edilmemiş çözelti reaksiyonunda nükleofile olan iyi anlaşılmış kovalent bağ ile karşılaştırılmalıdır. Gerçek bir kovalent kataliz önerisi (bariyerin çözeltideki karşılık gelen engelden daha düşük olduğu durumlarda), örneğin, bir enzim grubu (örneğin, çok güçlü bir hidrojen bağı) ile geçiş durumuna kısmi bir kovalent bağ gerektirecektir, katalize önemli ölçüde katkıda bulunmaz.
Metal iyon katalizi
Aktif bölgedeki bir metal iyon, şarj stabilizasyonu ve kalkanlamayı koordine ederek katalize katılır. Bir metalin pozitif yükü nedeniyle, metal iyonları aracılığıyla yalnızca negatif yükler dengelenebilir.[22] Bununla birlikte, metal iyonları biyolojik katalizde avantajlıdır çünkü pH'daki değişikliklerden etkilenmezler.[23] Metal iyonları, bir Lewis asidi olarak hareket ederek suyu iyonize etme görevi de görebilir.[24] Metal iyonları ayrıca oksidasyon ve indirgeme ajanları olabilir.[25]
Bağ suşu
Bu, enzimin geçiş durumuna afinitesinin substratın kendisinden daha büyük olduğu indüklenmiş uygun bağlanmanın temel etkisidir. Bu, substrat bağlarını geçiş durumunun konformasyonuna daha yakın bir konuma zorlayan yapısal yeniden düzenlemeleri indükler, böylece substrat ile geçiş durumu arasındaki enerji farkını azaltır ve reaksiyonun katalize edilmesine yardımcı olur.
Bununla birlikte, gerinim etkisi, aslında, geçiş durumu dengeleme etkisinden ziyade bir temel durum dengesizleştirme etkisidir.[11][26] Dahası, enzimler çok esnektir ve büyük gerilme etkisi uygulayamazlar.[27]
Substrattaki bağ suşuna ek olarak, aktif bölgedeki kalıntıları aktive etmek için enzimin kendisi içinde bağ suşu da indüklenebilir.
| Örneğin: |
| Substrat, bağlı alt-tabaka ve geçiş durumu konformasyonları lizozim . |
| Substrat, bağlanma sırasında, heksoz halkasının yarım sandalye konformasyonundan (ekvator c6'yı eksenel pozisyonda olmaya zorlayan proteinin amino asitlerinin sterik engellemesi nedeniyle) bozulmuştur.[28] |
Kuantum tünelleme
Bu geleneksel "bariyer üstü" mekanizmalar, bazı durumlarda "bariyerden geçen" mekanizmaların (kuantum tünelleme) modelleri ve gözlemleriyle zorlanır. Bazı enzimler, klasik ΔG ‡ ile tahmin edilenden daha hızlı olan kinetiklerle çalışır. "Bariyerin içinden" modellerde, bir proton veya bir elektron aktivasyon engellerinden tünel açabilir.[29][30] Aromatik amin dehidrojenaz ile triptamin oksidasyonunda protonlar için kuantum tünelleme gözlenmiştir.[31]
Kuantum tünelleme büyük bir katalitik avantaj sağlamıyor gibi görünmektedir, çünkü tünelleme katkıları çözeltideki katalize edilmiş ve katalize edilmemiş reaksiyonlarda birbirine benzer.[30][32][33][34] Bununla birlikte, tünelleme katkısı (tipik olarak hız sabitlerini klasik 'bariyer üstü' rotası için reaksiyon hızına kıyasla ~ 1000 [31] kat artırma) biyolojik organizmaların yaşayabilirliği için çok önemlidir. Bu, biyolojide tünel oluşturma reaksiyonlarının genel önemini vurgulamaktadır.
1971-1972'de enzim katalizinin ilk kuantum mekanik modeli formüle edildi.[35][36]
Aktif enzim
Enzim-substrat kompleksinin bağlanma enerjisi, substrat aktivasyonu için gerekli olan harici bir enerji olarak kabul edilemez. Yüksek enerji içeriğine sahip enzim, ilk olarak enzimin katalitik bölgesinden birinci bağlı reaktantın son yerine bazı özel enerji grubu X1'i, ardından ikinci bağlı reaktanttan (veya tek reaktantın ikinci grubundan) başka bir X2 grubunu aktarabilir. Substratın ürüne dönüşümünü ve enzim rejenerasyonunu bitirmek için aktif bölgeye aktarılmalıdır.[37]
Tüm enzimatik reaksiyon, iki birleştirme reaksiyonu olarak sunulabilir:
-
(1)

-
(2)

Reaksiyon (1)'den görülebileceği gibi, aktif enzimin X1 grubu, hem elektrostatik inhibisyonu hem de atomların itilmesini önlemek için enzim içindeki değişim reaksiyonu olasılığı nedeniyle üründe görünür. Bu nedenle aktif enzim, enzimatik reaksiyonun güçlü bir reaktanı olarak temsil edilir. Reaksiyon (2), enzim içinde, grup X-2 kalıntıları için zeminin tamamlanmamış dönüşümünü göstermektedir. Fikir olarak bu yaklaşım daha önce varsayımsal son derece yüksek enzimatik dönüşümlere (katalitik olarak mükemmel enzim) dayanarak önerilmişti.[38]
Mevcut yaklaşımın doğrulanması için can alıcı nokta, katalizörün, reaksiyonun transfer grubu ile bir enzim kompleksi olması gerektiğidir. Bu kimyasal özellik, çeşitli enzimatik reaksiyonların iyi çalışılmış mekanizmalarıyla desteklenmektedir. Serin proteazlar ailesinin iyi çalışılmış bir üyesi olan saf bir protein α-kimotripsin (kofaktör olmadan hareket eden bir enzim) tarafından katalize edilen peptit bağı hidrolizinin reaksiyonu buna örnektir.[39]
Bu reaksiyon için deneysel sonuçlar iki kimyasal adım olarak sunulur:
-
(3)

-
(4)

Burada S1, bir polipeptit, P1 ve P2 ürünlerdir. Birinci kimyasal aşama (3), kovalent bir asil-enzim ara ürününün oluşumunu içerir. İkinci adım (4) deasilasyon adımıdır. Başlangıçta enzim üzerinde bulunan ancak suda bulunmayan H+ grubunun hidroliz aşamasından önce üründe göründüğüne dikkat etmek önemlidir, bu nedenle enzimatik reaksiyonun ek bir grubu olarak düşünülebilir.
Böylece reaksiyon (3), enzimin reaksiyonun güçlü bir reaktantı olarak davrandığını gösterir. Önerilen kavrama göre, enzimden lH taşıma (P1 ve P2 grupları arasında) birinci başlangıç kimyasal bağın birinci tepkime dönüşümü, sorun teşkil etmektedir. Hidroliz aşaması, ikinci kimyasal bağın bozulmasına ve enzimin yenilenmesine yol açar.
Önerilen kimyasal mekanizma, ortamdaki substratların veya ürünlerin konsantrasyonuna bağlı değildir. Bununla birlikte, konsantrasyonlarındaki bir kayma, esas olarak (1) ve (2) reaksiyonlarının ilk ve son aşamalarında, su çözeltisindeki S veya P olsun, her molekülün serbest enerji içeriğindeki değişiklikler nedeniyle serbest enerji değişikliklerine neden olur. Bu yaklaşım, kas kasılma mekanizmasına uygundur. İskelet kasında ATP hidrolizinin son adımı, miyozin başlarının aktin ile birleşmesinden kaynaklanan ürün salınımıdır.[40] Birleşme reaksiyonu sırasında aktin bağlama yarığının kapanması yapısal olarak miyozin aktif bölgesindeki nükleotit bağlama cebinin açılmasıyla birleştirilir.[41]
Özellikle, ATP hidrolizinin son aşamaları, hızlı fosfat salınımı ve ADP'nin yavaş salınmasını içerir.[42][43] Bağlı ADP anyonundan su çözeltisine bir fosfat anyonunun salınması, bir ekzergonik reaksiyon olarak düşünülebilir çünkü fosfat anyonu düşük moleküler kütleye sahiptir.
Böylece, inorganik fosfat H2PO4− 'nin birincil salınımının, ATP hidrolizinin serbest enerjisinin önemli bir kısmının, aktif akış üreten solvatlı fosfatın kinetik enerjisine dönüşmesine yol açtığı sonucuna varılır. Lokal bir mekanik-kimyasal transdüksiyon varsayımı, Tirosh'un kas kasılma mekanizmasıyla uyumludur; burada kas kuvveti, ATP hidrolizinin yarattığı entegre bir aktif akış eyleminden kaynaklanır.[44][45]
Katalitik mekanizma örnekleri
Gerçekte, çoğu enzim mekanizması birkaç farklı kataliz türünün bir kombinasyonunu içerir.
Trioz fosfat izomeraz
Trioz fosfat izomeraz iki trioz fosfat izomeri dihidroksiaseton fosfat ve D -gliseraldehit 3-fosfatın tersine çevrilebilir dönüşümünü katalize eder.
Enzim yayılımı
Tek moleküllü çalışmaların ortaya çıkışı, 2010'larda bağlanmamış enzimlerin hareketinin artan substrat konsantrasyonu ve artan reaksiyon entalpisi ile arttığı gözlemine yol açtı.[46] Daha sonraki gözlemler, difüzivitedeki bu artışın, enzimin kütle merkezinin geçici yer değiştirmesi ile tetiklendiğini ve bunun da "enzimi iten bir geri tepme etkisi" ile sonuçlandığını göstermektedir.[47]
Reaksiyon benzerliği
Enzimatik reaksiyonlar (EC) arasındaki benzerlik, bağ değişiklikleri, reaksiyon merkezleri veya alt yapı ölçütleri (EC-BLAST) kullanılarak hesaplanabilir.[48]
Ayrıca bakınız
Kaynakça
- Kamerlin (2010). "At the dawn of the 21st century: Is dynamics the missing link for understanding enzyme catalysis?". Proteins: Structure, Function, and Bioinformatics. 78 (6): 1339-75. doi:10.1002/prot.22654. PMC 2841229 $2. PMID 20099310.
- Srinivasan (27 Eylül 2020). "Words of advice: teaching enzyme kinetics". The FEBS Journal. doi:10.1111/febs.15537. ISSN 1742-464X.
- Koshland DE (Şubat 1958). "Application of a Theory of Enzyme Specificity to Protein Synthesis". Proc. Natl. Acad. Sci. U.S.A. 44 (2): 98-104. doi:10.1073/pnas.44.2.98. PMC 335371 $2. PMID 16590179.

- Modern Physical Organic Chemistry. University Science Books. 2006. ISBN 978-1-891389-31-3.
- Savir Y (2007). "Conformational Proofreading: The Impact of Conformational Changes on the Specificity of Molecular Recognition" (PDF). PLOS One. 2 (5): e468. doi:10.1371/journal.pone.0000468. PMC 1868595 $2. PMID 17520027. 14 Mayıs 2011 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 22 Ağustos 2010.
- Stanton (1998). "Combined ab initio and Free Energy Calculations To Study Reactions in Enzymes and Solution: Amide Hydrolysis in Trypsin and Aqueous Solution". J. Am. Chem. Soc. 120 (14): 3448-3457. doi:10.1021/ja972723x.
- Kuhn (2000). "QM-FE and Molecular Dynamics Calculations on Catechol O-Methyltransferase: Free Energy of Activation in the Enzyme and in Aqueous Solution and Regioselectivity of the Enzyme-Catalyzed Reaction". J. Am. Chem. Soc. 122 (11): 2586-2596. doi:10.1021/ja992218v.
- Bruice (1999). "Ground State and Transition State Contributions to the Rates of Intramolecular and Enzymatic Reactions". Acc. Chem. Res. 32 (2): 127-136. doi:10.1021/ar960131y.
- Page (1971). "Entropic Contributions to Rate Accelerations in Enzymic and Intramolecular Reactions and the Chelate Effect". Proc. Natl. Acad. Sci. USA. 68 (8): 1678-1683. doi:10.1073/pnas.68.8.1678. PMC 389269 $2. PMID 5288752.
- Warshel (2001). "Dynamics of Biochemical and Biophysical Reactions: Insight from Computer Simulations". Quart. Rev. Biophys. 34 (4): 563-679. doi:10.1017/s0033583501003730. PMID 11852595.
- Warshel (2006). "Electrostatic Basis of Enzyme Catalysis". Chem. Rev. 106 (8): 3210-3235. doi:10.1021/cr0503106. PMID 16895325.
- Warshel (1989). "How do Serine Proteases Really Work?". Biochemistry. 28 (9): 3629-37. doi:10.1021/bi00435a001. PMID 2665806.
- Biochemistry. John Wiley & Sons. cop. 2011. OCLC 808679090. Tarih değerini gözden geçirin:
|tarih=(yardım) - Marcus (1965). "On the Theory of Electron-Transfer Reactions. VI. Unified Treatment for Homogeneous and Electrode Reactions" (PDF). J. Chem. Phys. 43 (2): 679-701. doi:10.1063/1.1696792.
- Warshel (1978). "Energetics of Enzyme Catalysis". Proc. Natl. Acad. Sci. USA. 75 (11): 5250-5254. doi:10.1073/pnas.75.11.5250. PMC 392938 $2. PMID 281676.
- "How Enzymes Work"
- "EXTREME ELECTRIC FIELDS POWER CATALYSIS IN THE ACTIVE SITE OF KETOSTEROID ISOMERASE",
- Toney, M. D. "Reaction specificity in pyridoxal enzymes." Archives of biochemistry and biophysics (2005) 433: 279-287
- Micronutrient Information Center, Oregon State University
- Biochemistry. John Wiley & Sons Inc. 2004. ss. 986-989. ISBN 978-0-471-25090-6.
- Biochemistry. John Wiley & Sons Inc. 2004. ss. 604-606. ISBN 978-0-471-25090-6.
- Piccirilli (7 Ocak 1993). "Metal ion catalysis in the Tetrahymena ribozyme reaction". Nature (İngilizce). 361 (6407): 85-88. doi:10.1038/361085a0. PMID 8421499.
- "Metal Ion Catalysis of Nucleophilic Organic Reactions in Solution". Reactions of Coordinated Ligands. Advances in Chemistry. 37. AMERICAN CHEMICAL SOCIETY. 1 Ocak 1962. ss. 19-36. doi:10.1021/ba-1963-0037.ch002. ISBN 978-0841200388.
- Fife (1 Şubat 1985). "Divalent metal ion catalysis in the hydrolysis of esters of picolinic acid. Metal ion promoted hydroxide ion and water catalyzed reactions". Journal of the American Chemical Society. 107 (4): 1041-1047. doi:10.1021/ja00290a048. ISSN 0002-7863.
- Stadtman (1 Ocak 1990). "Metal ion-catalyzed oxidation of proteins: biochemical mechanism and biological consequences". Free Radical Biology & Medicine. 9 (4): 315-325. doi:10.1016/0891-5849(90)90006-5. ISSN 0891-5849. PMID 2283087.
- Catalysis in Chemistry and Enzymology. reprint. New York: Dover Publications. 1987 [1969]. ISBN 9780486654607.
- Warshel (1976). "Theoretical Studies of Enzymatic Reactions: Dielectric Electrostatic and Steric Stabilization of the Carbonium Ion in the Reaction of Lysozyme". Journal of Molecular Biology. 103 (2): 227-49. doi:10.1016/0022-2836(76)90311-9. PMID 985660.
- <fundamentals of biochemistry Voet, voet and Pratt 4th edition>, which is similar in shape to the transition state.
- Garcia-Viloca (2004). "How enzymes work: analysis by modern rate theory and computer simulations". Science. 303 (5655): 186-95. doi:10.1126/science.1088172. PMID 14716003.
- Olsson (2004). "Simulations of the large kinetic isotope effect and the temperature dependence of the hydrogen atom transfer in lipoxygenase". Journal of the American Chemical Society. 126 (9): 2820-8. doi:10.1021/ja037233l. PMID 14995199.
- Masgrau (2006). "Atomic description of an enzyme reaction dominated by proton tunneling". Science. 312 (5771): 237-41. doi:10.1126/science.1126002. PMID 16614214.
- Hwang (1996). "How important are quantum mechanical nuclear motions in enzyme catalysis". J. Am. Chem. Soc. 118 (47): 11745-11751. doi:10.1021/ja962007f.
- Ball (2004). "Enzymes: By chance, or by design?". Nature. 431 (7007): 396-397. doi:10.1038/431396a. PMID 15385982.
- Olsson (2006). "Dynamical Contributions to Enzyme Catalysis: Critical Tests of A Popular Hypothesis". Chem. Rev. 106 (5): 1737-1756. doi:10.1021/cr040427e. PMID 16683752.
- Volkenshtein M.V., Dogonadze R.R., Madumarov A.K., Urushadze Z.D., Kharkats Yu.I. Theory of Enzyme Catalysis.- Molekuliarnaya Biologia, Moscow, 6, 1972, 431-439
- Volkenshtein M.V., Dogonadze R.R., Madumarov A.K., Urushadze Z.D., Kharkats Yu.I. Electronic and Conformational Interactions in Enzyme Catalysis. In: E.L. Andronikashvili (Ed.), Konformatsionnie Izmenenia Biopolimerov v Rastvorakh, Publishing House "Nauka", Moscow, 1973, 153-157
- Foigel (2011). "Is the enzyme a powerful reactant of the biochemical reaction?". Molecular and Cellular Biochemistry. 352 (1–2): 87-9. doi:10.1007/s11010-011-0742-4. PMID 21318350.
- Fogel (1982). "Cooperativity of enzymatic reactions and molecular aspects of energy transduction". Mol. Cell. Biochem. 47 (1): 59-64. doi:10.1007/bf00241567. PMID 7132966.
- Hengge (2004). "Role of protein conformational mobility in enzyme catalysis: acylation of alpha-chymotrypsin by specific peptide substrates". Biochemistry. 43 (3): 742-747. doi:10.1021/bi030222k. PMID 14730979.
- Lymn (1971). "Mechanism of adenosine triphosphate hydrolysis by actomyosin". Biochemistry. 10 (25): 4617-4624. doi:10.1021/bi00801a004. PMID 4258719.
- Holmes (2003). "Electron cryo-microscopy shows how strong binding of myosin to actin releases nucleotide". Nature. 425 (6956): 423-427. doi:10.1038/nature02005. PMID 14508495.
- Siemankowski (1985). "ADP dissociation from actomyosin subfragment 1 is sufficiently slow to limit the unloaded shortening velocity in vertebrate muscle". Proc. Natl. Acad. Sci. USA. 82 (3): 658-662. doi:10.1073/pnas.82.3.658. PMC 397104 $2. PMID 3871943.
- White (1997). "Kinetics of nucleoside triphosphate cleavage and phosphate release steps by associated rabbit skeletal actomyosin, measured using a novel fluorescent probe for phosphate". Biochemistry. 36 (39): 11828-11836. doi:10.1021/bi970540h. PMID 9305974.
- Tirosh (1990). "Translational motion of actin filaments in the presence of heavy meromyosin and MgATP as measured by Doppler broadening of laser light scattering". Biochim. Biophys. Acta. 1037 (3): 274-280. doi:10.1016/0167-4838(90)90025-b. PMID 2178685.
- Tirosh (2006). "Ballistic protons and microwave-induced water solutions (solitons) in bioenergetic transformations". Int. J. Mol. Sci. 7 (9): 320-345. doi:10.3390/i7090320.
- Muddana, Hari S. (28 Ocak 2010). "Substrate Catalysis Enhances Single-Enzyme Diffusion". Journal of the American Chemical Society. 132 (7): 2110-1. doi:10.1021/ja908773a. PMC 2832858 $2. PMID 20108965.

- Riedel, Clement (8 Ocak 2015). "The heat released during catalytic turnover enhances the diffusion of an enzyme". Nature. 517 (7533): 227-30. doi:10.1038/nature14043. PMC 4363105 $2. PMID 25487146. Diğer özet – Nature: News & Views (8 Ocak 2015).
- Rahman (2014). "EC-BLAST: a tool to automatically search and compare enzyme reactions". Nature Methods. 11 (2): 171-174. doi:10.1038/nmeth.2803. PMC 4122987 $2. PMID 24412978.
Daha fazla okuma
- Alan Fersht, Protein Biliminde Yapı ve Mekanizma : Enzim Katalizi ve Protein Katlama Rehberi. WH Freeman, 1998.0-7167-3268-8ISBN 0-7167-3268-8