Böcek uçuşu
Böcekler omurgasızların tek grubudur ve bu grubun kanatları ve uçuş özellikleri gelişmiştir. İki böcek grubu, kız böcekleri ve mayıs sineği, kanatlarına eklenmiş uçuş kaslarına sahiptirler. Diğer böceklerde ise kaslar göğüs kısmına eklenmiştir, bocalamaya ve indirekt olarak kanatları vurmaya sebep olmaktadır. Bazı çok küçük böcekler aerodinamiklerini kararlı durumda tutamazlar fakat Weis-Fogh hafifçe vuruşu ve fırlama mekanizmasından, üretilen yüksek kaldırma kuvveti kanatların yırtılmasına hırpalanmasına sebep oluyor. Birçok böcek havada durabilir, devamlı yükseklikte ve konum kontolü ile. Bazı böcekler güve gibi arka kanatlarının uyumu için ön kanat çiftlerine sahiptir.

Böceklerin ilk uçuşu karbonifer olarak bilinen jeolojik zaman diliminde gerçekleşmiştir, yaklaşık 350 milyon yıl önceye denk gelmektedir. Kanatlar yanlardaki uzuvlarına ek olarak gelişmiştir, diğer amaçları için zaten sinirlere, eklemlere ve kaslara sahiplerdir. Bunlar başlangıçta belki de denize açılma için kullanılıyordu veya süzülürken iniş hızını azaltmak için kullanılıyordu.
Mekanizma
Direkt Uçuş


Diğer böceklerin aksine, özel kız böceklerinin (yusufçuk ve küçük kız böceği) kanat kasları ( geleneksel düzende bir tür "ilkin böcekleri" olarak tanımlanır.) temel olan kanatlarına üzerine direkt eklenir, aşağı doğru küçük hareketler için olan bu kanatlar hareketlidir yani açılır kapanırdır, yukarı kaldıran kanatlarda kendi kendine yukarı doğrudur, havada kürek çekmeye çok benzer yusufçuklar ve küçük kız böcekleri benzer şekilde ve büyüklükte ön ve arka kanatlara sahiptirler ve bağımsız olarak çalışırlar, bu onlara iyi kontrol derecesi ve yönlerini ve hızlarını değiştirmeleriyle birlikte hareketlilik açısından tutarsızlık verir ve diğer uçuşan böcekler görülmez. Tabii ki bu sürpriz değildir, verilen kız böceklerinin tamamı hava yırtıcıdır ve daima diğer hava yırtıcı böcekleri avlarlar .[1]
Indirekt uçuş
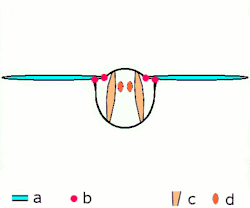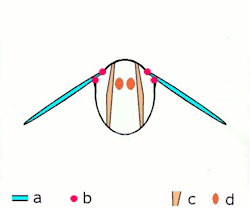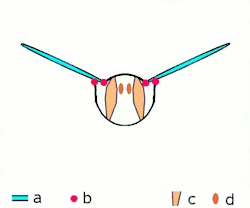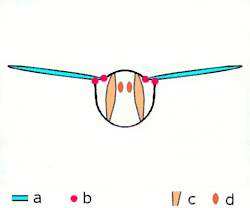

Direkt uçuş kaslarına ek olarak iki düzenleri vardır, diğer tüm yaşayan kanatlı böcekler uçarken farklı mekanizmalar kullanırlar, endirekt uçuş kasları da dâhil. Bu mekanizma bir zamanlar gelişime uğramıştır ve bu bir özellik olarak tanımlanır biyolojik sınıflamada gelişmiş böcekler sınıfına girer; bu şuna karşılık olarak gelir, muhtemelen tesadüfen değildir, kanat katlama mekanizmasının görünüşü ile birlikte, bu gelişmiş böceklere dinlenirken kanatlarının karınlarına doğru çekilmesine izin verir. (bu yeteneğe karşın ikincil bazı gruplarda olmayan bir özelliktir, mesela kelebeklerde).[1]
İki fonksiyonal kanat çiftleriyle beraber yüksek gruptalardır, bu çiftler birçok yolla mekanik olarak birliktedirler ve bu birçok ilkel grup için yanlış olmasına rağmen, fonksiyolanlar tek kanat olarak bulunurlar. Ayrıca gelişmiş böcekler arasından istisnalar vardır; hayalet güveler kanatların çiftlerini açabilirler ve bağımsız olarak hareket edebilirler, yusufçuklar gibi üzerlerinde uçabilirler. Tüm yusufçuklar ne paylaşırsa, fakat karınlarındaki kasların çalışmasının bir yoludur: bu kaslar, kanatların eklenmesinden ziyade, karın bölgesine eklenmesi ve deforme olmasıdır; çünkü kanatlar torasik dış iskeletin genişlemesi, karın bölgesinin deformasyonu kanatların hareketine de sebep olur. Boyuna uzayan sırt kaslarının oluşumunda sırtttan karna olan baskısı, kanatları aşağı çevirerek karnın arka yüzeyinin yukarı yaylanmasına sebep olur. Tergosternal kasların oluşumunda göğüs kısmının arka tarafı yine aşağı doğru çekilir, kanatların aşağı çekilmesine sebep olur. Bazı gruplarda, tergosternal kaslar gevşediğinde, aşağı vuruş sadece karnın geri çekilmesinde başarılıdır. Birçok küçük sklerite kanat temellerinde var, ayrılmış, kaslar eklenmişti ve bunlar kanat kontrolünün iyi olmasında kullanılır ve bu yollar eğimde birçok ayarlamaya ve kanat çırpış genişliklerine izin verir.Son düzeltmelerden biri de şudur gelişmiş canlılarırn görünüşleri (kınkanatlar, sinek ve zar kanatlılar) kaslı ve sinirsel sistemleri olan canlılara örnektir mademki bir sinir dürtüsü kas liflerinin birçok kasılmasına sebep olur ; bu kanat vuruşu oranın artması için frekansa izin verir ve bu sinir sistemlerine uyarının gönderilmesine sebep olur. Bu özelleştirilmiş kasların formu belirlenmiş asenkron uçuş kasıdır. Tüm etki birçok gelişmiş kanatların vuruşu böcek direkt uçuş kaslarından hızlıdır.[1]
Aerodinamikleri
Böcek uçuşlarının iki temel aerodinamik modeli vardır: yaratılan, öncü girdap .
Öncü Girdap
Birçok böcek metot kullanarak helezonik uçuş kenarı girdabı yaratır. Bu kanat çırpma hareketleri basit yarım çırpışlar boyunca olur. Aşağı yönde yapılan iniş başlar ve biter ve bu aşağı doğru ve ileri doğru dalınır. Ön cepheyi arka tarafa sabitlemek amacıyla kanatlar çabucak ters çevrilir. Yukarı vuruş ayrıca kanatların yukarı ve arkaya itilmesidir. Bunun üzerine kanatlar tekrar ters dönmüştür (dolaylama) ve aşağı yönlü vuruş oluşur. Böceklerde frekans aralığı eş zamanlı frekans uçuş kasları genel olarak 5 ila 200 hertz (Hz) arasındadır. Bunların içinde eş zamanlı olmayan kaslar da vardır, bu durumda kanat vuruş frekansı 1000 Hz. Böceklerin süzülüşü sırasında, iki vuruş aynı süre içerisinde tamamlanır. Kısa aşağı yönlü vuruş, fakat itme (akışkanlar mekaniğini). [2][3]
Temel kuvvetlerin tanımlanması böcek uçuşlarının anlamasında hassas bir konudur. Birinci deneme yarı kararlı kanat çırpışlarının tahmin edilmesi durumudur. Bunun anlamı hava herhangi bir zaman içerisinde kanatların üzerinde akar bunun öngörüsü ile kanat çırpımı olmaması, kanatların sabit durma durumundaki açıdaki akış ile aynıdır. Kanat çırpışları birçok hareketsiz konumda parçalara bölme yoluyla analiz edilebilir ve herhangi bir anda kanatların anlık kuvveti için zaman cetveli yaratılabilir. Bu hesaplanan kaldırma kuvveti üç katı küçük bulundu, böylece araştırmacılar bunun aerodinamik kuvvetini sağlayan düzensiz bir olay olduğunu fark etmişlerdir. Önceki zamanlarda ortalama uçuşun kanat çırpımına yakın olması için birçok geliştirici çözüm denenmiştir. Bazı araştırmacılar kuvvetin supinasyonda zirveye ulaşacağını tahmin etmişlerdir. Sirke sineği dinamik olarak pullanmıştır, bu kuvvet tahminleri daha sonra onaylanacaktır. Diğer tartışmalar supinasyon ve pronasyonun sebep olduğu bilinmeyen rotasyonel etki sırasında kuvvetin zirvede olduğunu söylemiştir. Bu rotasyonel etki temelde dönüşümsel olaylardan farklıdır. Bu yargıya karşı bazı zıt yargılar var. Hesaplamalı akışkanlar mekaniği boyunca, bazı araştırmacılar rotasyonal etki olmadığı konusunda tartışıyorlardı. Bu araştırmacılar yüksek kuvvetin bir önceki çarpıştaki dökülmenin sebebi olduğunu iddia ediyorlardı. [2][3]
Yukarıda bahsedilen rotasyonal etkiye benzer olarak, bu olay kanat çırpışlarının tam olarak anlaşılamaması ve üzerinde tam bir karar verilememesi ile ilgilidir. Çünkü bütün modeller tahmin edilebilir, farklı modellerde bu farzedilen etki hariç tutulur. Örneğin, Wagner etkisi şunları belirtiyor; akmazlıktan dolayı eğimli kanatlar hareketsizlikten ivmelenir. Bu olay kaldırma değeri ile açıklanabilir bu değer tahmin edilenden düşüktür. Tipik olarak, bu durumda havalanmanın eklenmesi ile kökenler bulundu. Bu etkinin akışkanlıkta Reynolds sayısı ile etkisiz olduğu çok önceden beri tartışılıyor bu tipik olarak bir böcek uçuşudur. Wagner etkisi etkisiz olarak değerlendirildi, bilinçli bir şekilde, en azından eski modelinde. Olaylardan en önemlisi ise uçuş sırasındaki ön tarafın emiş gücü ile oluşur. Bu kuvvet verimliliği hesaplamada çok önemlidir. Ön kenar emiş kavramı keskin kanatlı delta kanatlarında birinciyken girdap asansöründe dördüncüye konuldu. Saldırıların yüksek açılarda olanlarında, akış önde gelen kenarların üstünde ayrılır, tekrar eklenme ise arka kenara ulaşılmadan önce gerçekleşir. Ayrılmış akışların içindeki kabarcıklar girdaplardır. Hücumun saptırılması çok büyük olduğu için, akışın içine doğru birçok momentum gönderilir. Bu iki özellik büyük miktarda kaldırma kuvveti yaratır ve ayrıca biraz da ekstra direnç sağlar. Bu önemli özellik, ancak, kaldırmadır. Çünkü akış dağılmıştır, fakat hala büyük miktarda kaldırma kuvveti sağlar, bu olay perdövites olarak adlandırılır. Bu etki uçan böceklerin savruluşunda gözlemlendi ve bu it was proven to be capable of providing enough lift to account for the deficiency in the quasi-steady-state models. Bu etki kanoda birlikte kürek çekmede kullanılır.[2][3]
Kanat çırpmanın bütün etkileri aerodinamik olayların üç temel kökeniyle azaltılabilir: öncü girdap, kanattaki denge durumu aerodinamik kuvvetleri ve kanatların önceki çarpışın canlandırılmasındaki temastır. Uçan böceklerin büyüklük aralıkları yaklaşık olarak 20 mikrogramdan 3 grama kadar. Böceklerin vücut kütleleri artarken, kanat alanları genişler ve kanat vuruş frekansları azalır. Büyük böcekler için, Reynolds numaraları (Re) 10000 kadar büyük olabilir. Küçük böcekler için, bu 10 kadar az olabilir. Bunun anlamı akmazlık etkileri küçük böcekler için çok önemlidir, akışı tabakalı olmasına rağmen, hatta büyük uçuşlar için .[3][4]
Böceklerin vücudundaki diğer ilginç özellik ise vücut eğilimidir. Uçuş hızı artarken, böcek vücudu baş aşağı o eğilime yönelir ve daha çok yatay olur. Bu ön alanda azalma ve sonuç olarak vücut sürtünmesine sebep olur. Sürtünmenin artması hızın artması ile artar ve böcekler verimlilik daha önemliymiş gibi uçuşlarını daha verimli kılarlar. Ek olarak, aşağı yönlü vuruşlu saldırıların geometrik olarak açısal değişimleri aracılığıyla, mümkün olduğu kadar bütün manevraları süresince uçuşlarını maksimum verimlilikte tutarlar. Genel itme kuvvetinin gelişimi kaldırma kuvvetiyle kıyaslandığında oldukça küçüktür. Kaldırma kuvveti böceğin ağırlığından belki de üç kat fazladır, ama itme kuvvetinin en yüksek hızı vücut ağırlığının %20 sinden az hıza sahiptir. Bu kuvvet kanat çırpış hareketinin yukarı doğru daha kuvvetsiz olmasının temelidir.[3][5]
Kanat çırpışı ve savuruşu
Uçuşun ikinci metodu ise, çırpış ve savuruş veya Weis-Fogh mekanizması, fonksiyonlar farklıdır. Bu işlemde, küçük böcekler bulunur mesela trips ve chalcid eşek arısı, böcek vücudunun üzerinde kanatlar çırpılır ve daha sonra savuruş yapılır. Böcekler açık uçarlarken, hava emilir ve her kanadın üzerinde girdap oluşur. Bu girdap sınırı kanatların karşısına ilerler ve çırpış, diğer kanatlardaki girdabın oluşmasında rol oynar. Bu etki sayesinde, döngü ve böylece, kaldırma büyüklüğü genişlemek için artar, birçok durumda, tipik öncü girdap etkisinden büyüktür. Bu metodun ilk sebebi birçok böcek tarafından kaçınılmaz zararlarla ve kanatın giyilmesinin yol açılmasının oluşturulduğu tekrarlı çırpışın çalıştırılabilmesidir. Bu yaygındır, fakat böcekler arasında çok küçük ve düşük deneyimli Reynold numaralarıdır. Bu mekanizma ayrıca denize ait yumuşakçalar Limacina helicina, deniz kelebeği tarafından çalışıyor[6][7]
Temel denge
Sıvıdaki kanat hareketleri akışkanlar dinamiği tarafından deneyimlenir, bunu takip eden teamüller aerodinamikte bulunur, kuvvet bileşkeleri akış alan uzaklığına diktir ve kaldırma olarak tanımlanır (L), ve kuvvet bileşkeleri sürtünme akışkanlığıyla ters yönlüdür.(D). Reynold numaraları bu arada düşünüldü, uygun kuvvet birimi 1/2(ρU2S), ρ burada sıvı yoğunluğudur, S kanat alanı,ve U kanat süratidir. Ölçüsüz( boyutsuz) kuvvet kaldırma (CL) ve sürükleme (CD) katsayıları olarak adlandırılır, bu şu demek:[2]

CL ve CD sadece akış düzenli ise sabittir. Objelerin özel sınıfları mesela kanat profili sıvı boyunca saldırılar küçük açılarla parçalara ayrılırsa sabit duruma ulaşabilir. Bu durumda, kanat profili resim penetrasyon sınır şartlarının potensiyel akışla yatıştırılması aracılığıyla etrafında vizkos olmayan akış yaklaştırılabilir. Kutta-Joukowski teoremi ile kanadın 2D de daha öteye gidişi farz edilir ve bu şöyle belirtilir akış keskin kenarları pürüzsüzce akar, ve bu kanat profili etrafındaki toplam dönmeyi tanımlar. Bu kaldırma Bernoulli ilkesi (Blasius teoremi) ne karşılık gelmektedir:[2]

Kuşların ve böceklerin etrafındaki akış sıkıştırılamaz olarak düşünülür : Mach sayısı veya havada hareket ederken hız, tipik olarak 1/300 dür ve kanat frekansları yaklaşık olarak 10–103 Hz dir. Temel denklem olarak Navier-Stokes denklemleri kullanılır, kaymaz sınır şartları bulunur, denklem şudur:[2]

U(x, t) ın bulunduğu bölge akış alanıdır, p basınç, ρ sıvı yoğunluğudur, ν ise kinematik akmazlıktır, ubd ise sınırların hızıdır ve us ise katıların hızıdır. Uzunluk ölçeği seçilerek, L ve hız ölçeği, U, denklemler reynold numaralarını içeren boyutsuz formda açıklanabilirler. Re=UL/ν . Böcek kanatları ve kanat profili arasında iki temel farklılık vardır: Böcek kanadı daha küçüktür ve çırpılabilir. Yusufçukların kullanımları örnek olabilir, kanat genişlikleri (c) yaklaşık 1 cm, kanat uzunlukları (l) yaklaiık 4 cm ve kanat frekansları yaklaşık 40 Hz dir. Uç hızları (u) yaklaşık olarak 1 m/s dir ve reynolds numaralarına karşılık gelir, Re=uc/ν yaklaşık103. Sonuna küçülerek gider, a Chalcid yabanarısı yaklaşık 0,5–0,7 mm kanat uzunluğuna sahiptir ve kanat vuruşları yaklaşık 400 Hz dir. Reynold numaraları yaklaşık 25 tir tir. Reynold numaraları dizisi böcek uçuşları için hemen hemen 10 ila 104 arasındadır, bu iki sınırın teori için dönüşümüdür: yüzen bakteriler aracılığıyla kanat profili boyunca sürtünmesiz sabit akış ve vuruş akışı deneyimlenir. Bu yüzden, bu ortalama dizi tam olarak anlaşılamamıştır. Diğer yandan, bu belki de bizim gördüğümüz şeyler arasında en fazla her yerde birden bulunabilecek bir rejimdir Düşen yapraklar ve tohumlar, balıklar ve kuşlar hepsi düzenli olmayan akışta tesadiftür tıpkı böceklerde görüldüğü gibi.[2]
Reynold numaralarına ek olarak, en az iki diğer ilgili boyutsuz parametre vardır. Kanat üç hız ölçeğine sahiptir: vücuda göre çırpma hızı (u), vücudun ön hızı (U0), ortalama düşüş hızı (Ωc). Bunların oranları iki çeşit boyutsuz da oluşur,U0/u ve Ωc/u, önceki genellikle gelişmiş oran olarak tasvir edilir,ve bu ayrıca azaltılmış frekansla ilgilidir, fc/U0.[2]
Böcek kanatları sert ise, örneğin, Drosophila kanadı yakındır böylece, bunun hareketleri sabitlenmiş vücutla ilgilidir ve üç çeşitle tanımlanabilir: küresel koordinat sisteminin uç konumları, (Θ(t),Φ(t)), ve yükselme açısı ψ(t), eksen bağlanımı kök ve uçtur. Aerodinamik kuvveti tahmin edebilmek için pala elementi analizinden yola çıkılır, hücum açısını tanımlamak için bu ayrıca gereklidir (α). Tipik hücum açısı 70% kanat genişliği sıralaması süzülen böceklerde 25°den 45° dir.(sinekkuşunda 15°). Verinin varlığı birçok böcekte uygun olmasına rağmen, oranla daha az deney çarpış sırasındaki vuruş oranını rapor etmiştir. They include wind tunnel experiments of a tethered locust and a tethered fly, and free hovering flight of a fruit fly.[2]
Çünkü bunlar nispeten kolay ölçülebilir, kanat ucu yörüngeleri daha sıklıkla rapor edildi. Örneğin, seçilen tek uçuş ardışıklığı ağırlığı sağlayabilmek için yeterli kaldırma üretilir, bu şunu gösterir ki kanat ucueliptic bir şekil izler. Çapraz olmayan şekiller diğer böcekler tarafından ayrıca rapor edilmiş. Kesin şekilleri önemsizdir, takılan aşağı doğru hareket şunu gösteriyor böcekler ağırlıklarına destek için kullanılan kaldırmaya ek olarak aerodinamik sürtünme kullanabilir .[2]
Havada durabilme
| Wing beats[8] | |
|---|---|
| Honeybee | 250 beats/s |
| Housefly | 190 beats/s |
| Bumblebee | 130 beats/s |
| Syrphid hoverfly | 120 beats/s |
| Hornet | 100 beats/s |
| Horsefly | 96 beats/s |
| Hummingbird hawkmoth | 85 beats/s |
| Aeschnid dragonfly | 38 beats/s |
| Scorpion fly | 28 beats/s |
| Damselfly | 16 beats/s |
| Large white butterfly | 12 beats/s |
| Wing speed[8] | |
|---|---|
| Aeschnid dragonfly | 15.6 mph |
| Hornet | 12.8 mph |
| Hummingbird moth | 11.1 mph |
| Horsefly | 8.8 mph |
| Syrphid hoverfly | 7.8 mph |
| Bumblebee | 6.4 mph |
| Honeybee | 5.7 mph |
| Housefly | 4.4 mph |
| Damselfly | 3.3 mph |
| Scorpion fly | 1.1 mph |
Birçok böcekler havada durabilir veya havada bir noktada durabilirler, kanatlarını ani olarak vurarak bunu yapabilirler. Bunu yapabilme yeteneği, karşın, bu komplekstir; yerçekimi kuvvetine karşı gelebilmek için gerekli kaldırma kuvvete ulaşmak ve iki taraflı sabitliğin kullanılması gereklidir. Kaldırma kuvveti kanatların aşağı vuruşula oluşur. Hava ile çevrelenmiş bir bölgede kanatlar aşağı doğru itilmiştir, sonuç olarak havanın kanatlar üzerindeki reaksiyon kuvveti böcekler de yukarı doğrudur. Birçok böceğin kanatları gelişmiştir ve sonuç olarak yukarı doğru kanatlarını vurduğu sırada kanatların daki kuvvet küçük olur.Ölçümün ilk vuruş kuvveti ve dengesiz vuruş kuvvetinden dolayı böcekler sırasıyla aşağı yukarı,böcek salınımı ve kanat yukarı pozisyonunda sabit olarak dururlar.[4]
Kanat vuruşları böcek düşüşleri arasındaki uzaklık ani kanat vuruşlarına bağlıdır. Düşük bir oranla kanat vuruşları olursa, kaldırma kuvvetinin artık sıfır olduğu zaman aralığında ve sonuç olarak böcek düşüşleri eğer kanatlarını daha hızlı çarparlarsa daha uzağa olur. Böcekler verilen genlik stabilizelerine devam etmek için kanat çarpış frekansları gereken bir işlem yapılabilir. Hesaplamaları basitleştirmek için, bir farz ediliş vardır bu şudur kanatlar aşağı doğru giderken kaldırma kuvveti sonlu sabit bir değerdir, yukarı doğru giderken ise sıfırdır. Yukarı kanat vuruşları zaman aralığı sırasında, yerçekiminin etkisiyle h uzaklığı kadar aşağı düşer. [4]

Yukarı vuruşun onarımından sonra böcekler kendi duruşuna döner. Tipik olarak, 0,1 mm den fazla olmayan böcek değişimleri dikey pozisyonlarda gereklidir (örneğin, h=0,1 mm).Serbest düşmedeki maksimum izin verilen zaman [4]

Süreç sırasında kanatların yukarı hareket ve aşağı hareketleri hemen hemen eşittir, dönem T yukarı aşağı kanat hareketi iki kere tekrarlandığında tamamlanır Δr, bu şudur ,[4]

Vuruşların frekansı, f, anlamı saniyedeki kanat vuruşudur, denklem şu şekilde gösterilir :[4]

Örneklerde frekanslar 110 vuruş/s olarak kullanıldı, bu böceklerde bulunan tipik frekanstır. Kelebekler çok daha yavaş frekansa sahiptirler yaklaşık 10 vuruş/s kadar ve bunun anlamı onlar havada duramazlar. Diğer böcekler 1000 vuruş/s kadar frekans üretebilirler. Böcekleri eski haline getirmek için yani dikey pozisyonlarına getirmek için, aşağı vuruş sırasında, ortalama kuvvet, ortalama yukarı gücü böceklerin ağırlığının yaklaşık iki katı olmalıdır. Ve burası önemli çünkü böceğin üstüne uygulanan yarı zamanlıdır, ortalama yukarı kuvvet kütlesiyle basitleştirilmiştir .[4]
Güç girişi
Havada duruş için, gerekli olan gücü şimdi hesaplanabilir, böceklerin kütleleri 0,1 g tekrar düşünülerek, ortalama kuvvet, ortalama kuvvet, aşağı vuruş sırasında kanatlar tarafından uygulanır ve ağırlığın iki katı kadardır. Çünkü kanatlar tarafından uygulanan basınç, kanadın tüm yüzeyi üzerinde eşit bir biçimde dağılmış durumdadır, bunun anlamı her kanattaki yalnız bir noktanın kanadın orta noktasındayken üretilen kuvvet olduğunu farz edelim. Aşağı vuruş sırasında, kanatların merkezi dikey uzaklıkta d her iki yönde hareket eder. Böcekler tarafından her aşağı vuruştaki kuvvetin ve uzaklığını çarpımı toplam işi verir; bu şudur,

Eğer kanatlar her vuruşta 70° ile sallanırsa, n-bu durum şunu gösteriyor 1 cm uzunluğunda kanadı olan ve uzaklığı 0.57 cm olan böcekler içindir. Sonuç olarak her vuruşta iki kanat aracılığıyla iş yapılır :[4]

Sonra, enerji başka bir yere gitmek zorundadır ; burada, kullanılan örnekte, böceğin kütlesi her aşağı vuruşta 0.1 mm artar . Enerji E bu durum için gereklidir:[4]

Bu toplam enerjinin önemsiz açık bir biçimde kullanılan kesir, enerjinin çoğu işlemde harcanmıştır. Problemin daha detaylı analizi şunu gösteriyor, öncelikle kanatlar aracılığıyla yapılan iş öncelikle havadaki kinetik enerjiye dönüşüyor ve bu kanatların aşağı yönlü vuruşları ile ivmeleniyor. Güç bir saniyede yapılan işin miktarıdır; böceklerdeki kullanılışı örnekteki, gibidir, saniyede 110 aşağı yönlü vuruş yapar. Sonuç olarak, güç verimi P, saniyedeki vuruş ve bunun anlamı onun güç verimi P :[4]

Güç çıkışı
Gücün hesaplanmasında havada duruşu kullanılır, örnekler de hareket eden kanadın kinetik enerjisi önemsiz kabul edilir. Böceklerin kanatları, onlar kadar hafiftir, sonlu kütleye sahiptirler; sonuç olarak, hareket ederlerken bir kinetik enerjiye sahiptirler. Çünkü kanatlar dönme hareketi yaparlar, her vuruş sırasındaki maksimum kinetik enerji :[4]

Burada I kanatların eylemsizlik döngüsüdür ve maksimum açısal hız ise kanat vuruşları sırasındaki maksimum açısal hızıdır. Kanatların eylemsizlik döngüsünü elde etmek için, biz farz ederiz ki kanatlar dönel çubukların birinin sonuna konulması aracılığıyla. Kanatların eylemsizlik döngüsü şudur :[4]

l kanatların uzunluğu ve (1 cm) m ise iki kanadın ağırlığıdır, bu tipik olarak 10−3 g. Maksimum açısal hız, ωmax, maksimum hız dan kanadın merkezine νmax hesaplandı. [4]

Her vuruş sırasında kanatların merkezi ortalama doğrusal bir hızla νav, merkezdeki kanatların ayrılmasıyla d uzaklık ile, Δt zaman aralığındaki kanat vuruşları ile hareket eder. Bizim eski örneğimizden, d=0.57 cm ve Δt=4,5×10−3 saniye. Sonuç olarak:[4]

Kanat çırpışların başında ve sonunda kanatların hızı sıfırdır, bunun anlamı maksimum doğrusal hız ortalama hızdan fazladır. Eğer hızın kanat yolu boyunca sinüzoidal bir şekilde çeşitlendiğini farz edersek, maksimum hız ortalama hızın iki katı kadar fazladır. Böylece, maksimum açısal hız şudur :[4]

Ve sonuç olarak kinetik enerji budur:[4]

Çünkü iki kanat çırpışı vardır(yukarı yönlü ve aşağı yönlü) kanat hareketinin her devrinde, kinetik enerji 2×43=86 erg. Bu hemen hemen havada dururken tüketilen enerji miktarı kadardır.[4]
Esneklik
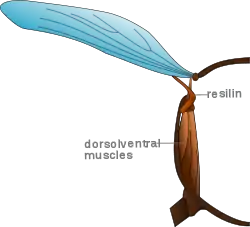
Böcekler kinetik enerji kazanır, ivmelenme olurken kaslar aracılığıyla sağlanır. Kanatlar vuruşun sonlarında azalarak ivmelenmeye başladığında, bu enerji dağılmak zorundadır. Aşağı vuruş sırasında, kaslar aracılığıyla kinetik enerji dağılır ve bu ısıya döner ( bu sıcaklık bazen vücudunun öz sıcaklığını korumasında kullanılır). Bazı böcekler kanatların yukarı hareketine yardım sağlamak için kinetik enerjiden fayda sağlayabilirler. Kanatlar böceklerin eklemleridir ve elastik ped ile protein gibi kauçuk içerir buna resilin denir. Kanatların yukarı vuruşu sırasında, resilin gerilir. Kanatlardaki kinetik enerji gerilen resilinde potansiyel enerjiye dönüşür ve bu yay gibi enerji depolar. Kanatlar aşağı doğru giderken, enerji bırakılır ve bu aşağı yönlü çırpışa yardım sağlar.[4]
Çok az basitleştirilmiş varsayım kullanarak, gerilmiş resilindeki depo edilen enerji miktarını hesaplayabiliriz. Resilin karşık şeklin içerisinde bükülmüş bir şekilde olmasına rağmen, örnekte verildiği gibi hesaplamalar düz çubuğun alanı ve uzunluğudur. Buna ek olarak, farz edelim ki resilinin gerilimi boyunca Hooke yasasına uyar. Bu tam olarak doğru değildir çünkü resilin düşünebilir miktarda gerilir ve sonuç olarak hem alan hem Young modülü gerilme işleminde değişir. Enerji E resilin de depolanır:[4]

Resilin için Y Young modülüdür, bu 1,8×107 dyn/cm2 olarak ölçüldü. Tipik olarak, böceklerin içinden arıların büyüklüğünde resilin hacmi silindir ile eşit 2×10−2 cm uzunluğunda ve 4×10−4 cm2 alanındadır. Bu örnekte verilmiştir, resilin çubuğunun gerilince uzunluğu %50 oranında artar. Bu şu demek, Δℓ yani uzunluk 10−2 cm.Sonuç olarak, bu durumda her bir kanatta resilin de enerji depo edilir :[4]

İki kanatta depolanan enerji 36 erg dir, bu yukarı vuruşlu kanatların kinetik enerjisiyle karşılaştırılabilir. Deneyler gösteriyor ki kanatlardaki kinetik enerjinin 80% i kadarı resilin de depo ediliyor.[4]
Kanat çiftleri
Bazı dört kanatlı böcek türlerinde, örneğin Lepidoptera, geniş alanda çeşitli morfolojil iki kanatlı mekanizmasında gelişimini tamamlamış olarak gelişti bu taksonları fonksiyon olarak "iki kanatlı”sunar. Hepsi, fakat çoğu temel formdaki, iki kanatlı olarak bulunur.[9]:4266
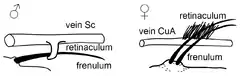
Mekanizmalar üç farklı tipte oluşur - jugal, frenulo-retinacular ve amplexiform.[10]
Genellikle ilkel gruplar genişlemiş temel arka planın yan alanı gibi lop a sahiptirler örneğin ön kanatların temeli jugum olarak adlandırılır bu uçarken arka kanatların altını kıvırır.[9][11]
Diğer gruplar arka kanatların üstünde fenuluma sahiptir ve bu ön kanatların altında retinakülüm asılıdır.[11]
Kelebeklerde ( hesperiid lerin bir tür erkeği hariç) ve Bombycoidealerde ( Sphingidae lar hariç), kanat çiftlerinde frenulumun ve retinakülümün bir sırası yoktur. Yerine, arka kanatların genişlemiş omuz alanı ön kanatlar aracılığıyla genişçe üst üste gelir. Özel mekanik bağlantı kaybı yerine, kanatlar aşama aşama üst üste düzenlenir. Ön kanatların güç vuruşu, ön kanatları uyumlu bir şekilde aşağı doğru iter. Bu tip çiftler frenate tipi çeşitliliktir fakat burada frenulum ve retinakülüm tamamen kaybolur .[12][13]
Biyokimya
Böcek uçuşlarında biyokimya kayda değer çalışmalara odaklanmışlardır. Birçok böcek karbonhidrat ve yağı uçuşta enerji kaynağı olarak kullanırken, birçok kın kanatlı ve sinekler enerji kaynağı olarak amino asit kullanırlar . Bazı türler iki türüde kullanırlar ve arı benzeri kelebekler mesela manduca sexta ısınma hareketi olarak ön uçuşta karbonhidrat kullanırlar. [14]
Kanat vuruşları bir saniyede 100 kereden az olan böcekler senkronize kaslarını kullanırlar. Senkronize kaslar bir kas tipidir ve bu her sinir sinyali için bir sözleşme gibidir, bu hızlı uçuşlar için çok verimlidir. Kanat vuruşları çok hızlı olan böcekler mesela bombus eş zamanlı olmayan kaslarını kullanırlar; bu kas türü bir sinir sinyalinden fazlası için olan bir sözleşme gibidir. Bu kaslar anlaşma için tarafından tekrar uyarılması ve kaslardaki gerilimin bırakılması aracılığıyla başarıya ulaşmıştır, bu basit tek başına uyarılma olayından daha hızlı gerçekleşir.[15]
Gelişim ve Adaptasyon

Karbonifer periyodunda bazı zamanlar, 350 milyon yıl önce ve bu zamanlarda sadece iki ana kütle bulunurdu, böcekler uçmaya başladı. Nasıl veya niçin böcek kanatları gelişmedi anlaşılamadı, alt karbonifer gelişimleri periyodunda büyük ölçüde uygun fosillerin kıtlığından dolayı büyük ölçüdedir. Böcek uçuşları kökeninde üç temel teori şunlar kanatlar paranotal loblardan gelişmiştir, toraks terganın gelişimi; bunlar taşınabilir karın modifikasyonlarıdır, suda yaşayan mayıs sineğinin nimfolarında bulunurlar veya toraks çıkıntılarından radyatörler kullanılarak gelişirler.[16]
Paranotal hipotezleri
Paranotal hipotezler şunları öne sürüyor böceklerin kanatları paranotal loblardan gelişti, adaptasyon öncesi bulunan böcek fosilleri şuna inandırıyor sekerken veya düşerken dengede tutulmak için destekte bulunuldu. Bu hipotezin ayrıcalığı çoğu böceğin eğilimidir, kolları tırmanırken ürkmek, zemine düşüşten kaçmak için. Bu tür loblar paraşüt olarak hizmet verdi ve böceklerin zemine yumuşak bir şekilde inmesine olanak sağladı. Teori şunu öneriyor bu loblar yavaş yavaş büyüyor ve daha sonraki aşamalarda göğüs kafesine ortak olacaktır. Hatta daha sonra bu ham kanatları hareket ettirmek için yeni kaslar belirecek. Bu model şunu gösteriyor kanatlardaki verimlilik kademeli olarak artacaktır, paraşütle başlayarak, sonra süzülerek gitme ve en son olarak uçuş aktif olacaktır. Hala, kanatların gelişimindeki fosil kanıtlarının temel eksikliğindeki eklemler ve kaslar bu teori için büyük sorun ortaya çıkartıyor, eklemliliğin ve damar düzeninin gelişimindeki anlık değişimler olarak görünüyor ve uzmanlar tarafından bu alanda bu durum reddediliyor.[16]
Epicoxal hipotezleri
Bazı entomolojistler şunu gösteriyor böcek kanatlarında muhtemel kökende su böceklerinde taşınabilir karın solungaçları bulundu, nimfa ve mayıs sineği örnek gösterilebilir. Bu teoriye göre bunlar soluk borusu solungaçları, onların yolları solunum sistemlerinin ve daha sonra düzenlenen hareket ettirici amaçlar ile var oluyor, sonunda kanatlarının içinin gelişimi için. Trakea solungaçları kanatçıklar ile donanımlıdır ve bunlar sürekli olarak titreşir ve küçük doğrusal kaslara sahiptirler.[16]
Endite-exite hipotezleri
Bu hipotez belki de en güçlü kanıta sahip hipotez ve şöyle diyor endites ve exites hangi adaptosyondan kaynaklanıyor, temel eklem bacaklılar arasında sırasıyla içerideki ve dışarıdaki uzantılardır. Trueman temel alınarak Goldschmidt aracılığıyla 1945'te Drosophila melanogaster üzerinde geliştirildi, in which a pod variation displayed a mutation transforming normal wings to what was interpreted as a triple-jointed leg arrangement with some additional appendages but lacking the tarsus, kanatların kostal yüzeyi normal olarak olabilir. Bu mutasyon sırt exite ve endite füzyonu için kuvvetli bir kanıt olarak tekrar yorumlanmıştı, bacaktan ziyade, uzantı ayarlaması ile daha iyi bir hipotez olmuştur. İnnervasyon, eklem ve kas kanatların gelişiminde gerekli ve zaten podomoreste vardır.[17]
Ayrıca bakınız
- Insect wing
- Unmanned aerial vehicles
- Insect migration
- Bird flight
- Flying and gliding animals
Notlar
- Chapman, A. D. (2006). Numbers of living species in Australia and the World. Canberra: Australian Biological Resources Study. ss. 60pp. ISBN 978-0-642-56850-2. 9 Haziran 2009 tarihinde kaynağından arşivlendi. Erişim tarihi: 23 Nisan 2016.
- Wang, Z. Jane (2005). "DISSECTING INSECT FLIGHT" (PDF). Annual Review of Fluid Mechanics. Cilt 37. Annual Reviews. ss. 183-210. Bibcode:2005AnRFM..37..183W. doi:10.1146/annurev.fluid.36.050802.121940. 16 Ocak 2014 tarihinde kaynağından (pdf) arşivlendi. Erişim tarihi: 23 Nisan 2016.
- Sane, Sanjay P. (2003). "The aerodynamics of insect flight" (PDF). The Journal of Experimental Biology. 206 (23). ss. 4191-4208. doi:10.1242/jeb.00663. PMID 14581590. 9 Temmuz 2011 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 23 Nisan 2016.
- Davidovits, Paul (2008). Physics in Biology and Medicine. Academic Press. ss. 78-79. ISBN 978-0-12-369411-9. 1 Ağustos 2020 tarihinde kaynağından arşivlendi. Erişim tarihi: 23 Nisan 2016.
- "Catching the Wake". Scientific American. 28 Haziran 1999. 18 Ekim 2012 tarihinde kaynağından arşivlendi. Erişim tarihi: 31 Mart 2011.
- Weisberger, Mindy (19 Şubat 2016). "Swim Like a Butterfly? Sea Snail 'Flies' Through Water". Scientific American. 31 Mart 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 20 Şubat 2016.
- Murphy, D., Adhikari, D., Webster, D., and Yen, J. (2016). "Underwater flight by the planktonic sea butterfly". Journal of Experimental Biology. Cilt 219. ss. 535-543.
- "Insect Wings in General". Aerodynamics of Insects. Cislunar Aerospace. 1997. 24 Mart 2011 tarihinde kaynağından arşivlendi. Erişim tarihi: 28 Mart 2011.
- Stocks, Ian.(2008).
- Scoble (1995) Subsec.
- Powell, Jerry A. Ch. Lepidoptera (pp. 631-664) in Resh, & Cardé (Eds).
- Dudley, Robert. (2002).
- Gorb, S. (2001) Ch 4.1.5 "Inter-locking of body parts".
- Joos, B. (1987). "Carbohydrate use in the flight muscles of Manduca sexta during pre-flight warm-up". Journal of Experimental Biology. Cilt 133. ss. 317-327.
- "Definition of Asynchronous muscle in the Entomologists' glossary". Department of Entomology, North Carolina State University. 13 Kasım 2015 tarihinde kaynağından arşivlendi. Erişim tarihi: 21 Mart 2011.
- Grimaldi, 1David; Engel, Michael S. (2005). Evolution of the Insects. New York, NY: Cambridge University Press.
- Trueman JWH (1990), Comment: evolution of insect wings: a limb exite plus endite model Canadian Journal of Zoology
- Thomas, Adrian L. R.; Norberg, R. Åke (1996). "Skimming the surface — the origin of flight in insects?". Trends in Ecology & Evolution. 11 (5). ss. 187-188. doi:10.1016/0169-5347(96)30022-0.
- Yanoviak, SP; Kaspari, M; Dudley, R (2009). "Gliding hexapods and the origins of insect aerial behaviour". Biol Lett. 5 (4). ss. 510-2. doi:10.1098/rsbl.2009.0029. PMC 2781901 $2. PMID 19324632.
Kaynakçalar
- Camper, M.A., "An Insect’s Role in the Development of Micro Air Vehicles," Colorado State University, 2003.
- Chapman, R.F. (1998) The Insects : Structure and Function. 4th Edition. Cambridge University Press.
- Dickinson, MH; Lehmann, FO; Sane, SP (1999). "Wing rotation and the aerodynamic basis of insect flight". Science. 284 (5422). ss. 1954-60. doi:10.1126/science.284.5422.1954. PMID 10373107.
- Ellington, Charles P.; Van Den Berg, Coen; Willmott, Alexander P.; Thomas, Adrian L. R. (1996). "Leading-edge vortices in insect flight". Nature. 384 (6610). ss. 626-630. Bibcode:1996Natur.384..626E. doi:10.1038/384626a0.
- Ellington, C. P. (1984). "The Aerodynamics of Hovering Insect Flight. I. The Quasi-Steady Analysis". Philosophical Transactions of the Royal Society B: Biological Sciences. 305 (1122). ss. 1-15. Bibcode:1984RSPTB.305....1E. doi:10.1098/rstb.1984.0049.
- Ellington, CP (1999). "The novel aerodynamics of insect flight: Applications to micro-air vehicles". The Journal of Experimental Biology. 202 (Pt 23). ss. 3439-48. PMID 10562527. 4 Mart 2016 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 26 Mayıs 2016.
- Grasmeyer, J.M. and Keennon, M.T., "Development of the Black Widow Micro Air Vehicle," AIAA Paper No. 2001-0127, 2001.
- Haj-Hariri, H., "Unsteady Aerodynamics of Flapping Wings," University of Virginia, 2001.
- Lewin, G. C.; Haj-Hariri, H. (2003). "Modelling thrust generation of a two-dimensional heaving airfoil in a viscous flow". Journal of Fluid Mechanics. Cilt 492. ss. 339-362. Bibcode:2003JFM...492..339L. doi:10.1017/S0022112003005743.
- Lighthill, M. J. (2006). "On the Weis-Fogh mechanism of lift generation". Journal of Fluid Mechanics. Cilt 60. s. 1. Bibcode:1973JFM....60....1L. doi:10.1017/S0022112073000017.
- Platzer, Max F., Department Chairman, "Aerodynamics and Aeroelasticity: Flapping-Wing Propulsion." Last revised 2005, Retrieved 1 November 2005, from https://web.archive.org/web/20051101200423/http://www.aa.nps.navy.mil/programs/aero/propulsion/
- Polhamus, E.C., "A Concept of the Vortex Lift of Sharp-Edge Delta Wings Based on a Leading-Edge-Suction Analogy," Langley Research Center, 1966.
- Pringle, J.W.S., "Insect flight," Oxford Biology Readers, Vol. 52, 1975.
- Sane, SP (2003). "The aerodynamics of insect flight". The Journal of Experimental Biology. 206 (Pt 23). ss. 4191-208. doi:10.1242/jeb.00663. PMID 14581590.
- Savage, S.B.; Newman, B.G.; Wong, D.T.M. (1979). "The role of vortices and unsteady effects during the hovering flight of dragon flies". The Journal of Experimental Biology. 83 (1). ss. 59-77. 8 Ağustos 2016 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 26 Mayıs 2016.
- Smyth, T., Jr. 1985. Muscle systems, pp. 227–286. In M.S. Blum [ed.], Fundamentals of insect physiology. John Wiley and Sons, New York.
- Sotavalta, O (1953). "Recordings of high wing-stroke and thoracic vibration frequency in some midges". Biol. Bull. Woods Hole. Cilt 104. ss. 439-444. doi:10.2307/1538496.
- Van Den Berg, C.; Ellington, C. P. (1997). "The vortex wake of a 'hovering' model hawkmoth". Philosophical Transactions of the Royal Society B: Biological Sciences. 352 (1351). ss. 317-328. doi:10.1098/rstb.1997.0023. PMC 1691928 $2.
- Walker, J.A. (Eylül 2002). "Rotational lift: something difference or more of the same?". The Journal of Experimental Biology. 205 (Pt 24). ss. 3783-3792. PMID 12432002. 8 Ağustos 2016 tarihinde kaynağından (PDF) arşivlendi. Erişim tarihi: 26 Mayıs 2016.
- Walker, P.B., "Growth of circulation about a wing and an apparatus for measuring fluid motion," Reports and Mem., Aeronaut. Res. Com. no. 1402, 1931.
- Zbikowski, R. (2002). "On aerodynamic modelling of an insect-like flapping wing in hover for micro air vehicles". Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences. 360 (1791). ss. 273-290. Bibcode:2002RSPTA.360..273Z. doi:10.1098/rsta.2001.0930.
Dış bağlantılar
- An Insect’s Role In The Development Of Micro Air Vehicles
- Insect-like Flapping-Wing Micro Air Vehicles 4 Ocak 2006 tarihinde Wayback Machine sitesinde arşivlendi.
- The Novel Aerodynamics Of Insect Flight: Applications To Micro-Air Vehicles 4 Mart 2011 tarihinde Wayback Machine sitesinde arşivlendi.
- The aerodynamics of insect flight 9 Temmuz 2011 tarihinde Wayback Machine sitesinde arşivlendi.
- Flight muscles22 Ağustos 2004 tarihinde Wayback Machine sitesinde arşivlendi.
- Flow visualization of butterfly aerodynamic mechanisms
- The Flight Of The Bumblebee 4 Mart 2016 tarihinde Wayback Machine sitesinde arşivlendi.
- Insect Flight4 Mart 2006 tarihinde Wayback Machine sitesinde arşivlendi.
- T. J. Dean (1 Mayıs 2003). "Chapter 1 — Fastest Flyer". Book of Insect Records. University of Florida. 21 Mayıs 2016 tarihinde kaynağından arşivlendi. Erişim tarihi: 23 Nisan 2016.

{kind=link}
{kind=link}