Deaminasyon
Deaminasyon bir molekülden bir amin grubunun çıkarılması. Bu reaksiyonu katalizleyen enzimler deaminaz olarak adlandırılır.
İnsan vücudunda deaminasyon başlıca karaciğerde olur, ancak glutamat ayrıca böbrekler de deamine olur. Aşırı protein alındığında amino asitler deaminasyon yoluyla yıkıma uğrar. Amino asitten amino grubu çıkartıldıktan sonra amonyağa dönüşür. Amino asidin geriye kalanı başlıca karbon ve hidrojenden oluşur veya başka bir moleküle dönüştürülür veya enerji için oksitlenir. Amonyak aşırı miktarda hücrelere zehirli olduğu için üre döngüsü içinde enzimler ona karbondioksit ekleyerek onu üre veya ürik asite dönüştürürler. Üre ve ürik asit kana kolaylıkla geçip idrarla atılabilir.
DNA'da deaminasyon reaksiyonları
Sitozin
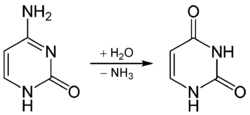
Spontan demainasyon, sitozin'in hidroliz yoluyla urasil'e dönüşmesi ve bu süreçte amonyak salmasıdır. DNA'da bu spontan (kendiliğinden olan) deaminasyon, meydana gelen urasil'in çıkartılması ile düzeltilir. Urasil normalde DNA'da bulunmayan bir baz olduğu için urasil-DNA glikosilaz enzimi tarafından çıkartılır. DNA zinciirnin o noktasındaki şeker grubunda bir baz takılı olmaması bir AP endonükleaz tarafından fark edilir, fosfodiester bağı orada kesilir. Meydana gelen hasar oraya bir diğer sitozin eklenmesiyle tamir edilir. A ailesi üyesi bir DNA polimeraz bu değişikliği yapar, bu işlem sırasında 3'-->5' eksonükleaz etkinliği ile DNA'da "çentik" olan yerden bir nükleotit çıkarır, sonra polimeraz etkinliği ile boşluğu gerekli olan nükleotit ile doldurur. DNA ligaz zincirdeki kopukluğu kapatmak için fosfodiester bağını yeniden oluşturur.
Deaminasyon reaksiyonu laboratuvar ortamında bisülfit kullanılarak da meydana getirilebilir. Bisülfit sitozin'in deamine olmasını sağlar ama 5-metilsitozin'e etki etmez. Bu özellikten yararlanılarak metillenmiş DNA'nın dizilemesinde metillenmemiş sitozin (urasil olarak belirir) ile metillenmiş sitozin (değişime uğramaz) ayırdedilebilir.
5-metilsitozin
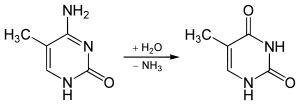
5-metilsitozin'in spontan deaminasyonu sonucu timin ve amonyak meydana gelir. Bu, en yaygın olan tek nükleotitli mutasyondur. DNA'da bu reaksiyon tamir edilemez çünkü tamir mekanizması timin bazını (urasil gibi) bir hata sonucu olmuş olarak algılayamaz. Eğer mutasyon bir genin fonksiyonunu etkileyip hücrenin (veya organizmanın) ölümüne yol açmazsa, varlığı devam eder. Tamir mekanizmasının bu yetersizliği, ökaryotik genomlarda CpG konumlarının ender olmasının nedenlerinden biridir.
Guanin
Guanin'in deaminasyonu sonucu ksantin oluşur. Ksantin, guanin'in enol totomerine benzer şekilde, sitozin yerine timin ile baz eşleşmesi yapar. Bunun sonucu, ikileşme sonrası br transisyon mutasyonudur, orijinal G-C baz çifti bir A-T baz çiftine dönüşür. Bu mutasyonun düzeltilmesi için baz çıkarma tamiri (baz eksizyon tamiri) sırasında bir alkiladenin glikosilaz enziminin katılımı gerekir.
Adenin
Adenin'in deaminasyonu sonucu hipoksantin oluşur. Hipoksantin, adenin'in imin totomerine benzer şekilde timin yerine sitozin ile baz eşleşmesi yapar. Bunun sonucu, ikileşme sonrası br transisyon mutasyonudur, orijinal A-T baz çifti bir G-C baz çiftine dönüşür
Bu işlemi yapan diğer enzimler
RNA biçimleme (RNA editing) enzimleri bazı mesajcı RNA moleküllerindeki spesifik bazları deamine ederek protein kodlayıcı dizilerini değiştirirler. Örneğin APOBEC geninin ürünü enzim, apolipoprotein B'nin mRNA'sında belli bir sitozin'i urasil'e dönüştürür. ADAR geni tarafında kodlanan Adenozin deaminaz, RNA-spesifik enzimi, çift iplikli RNA moleküllerinde adenozinleri inosin'e dönüştürür. Bazı tRNA moleküllerinde antikodon bölgesindeki bir Adenin inosin'e dönüştürülür.
