Arkea
Arkeler, Arkea (Yunanca αρχαία “eskiler”den türetme; tekil olarak Arkaeum, Arkaean veya Arkaeon) veya Arkebakteriler, canlı organizmaların bir ana bölümüdür.
| Arkea | ||
|---|---|---|
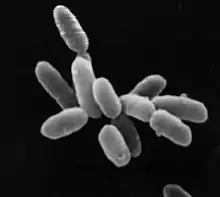 | ||
| Bilimsel sınıflandırma | ||
| ||
|
Crenarchaeota | ||
Yabancı literatürde bu gruptaki canlılar Archaea veya Archaebacteria, grubun tek bir üyesi ise tekil olarak Archaeum, Archaean veya Archaeon olarak adlandırılır
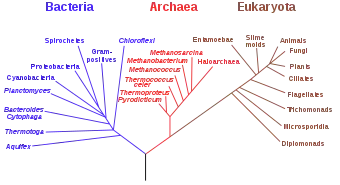
Arkeler, Bakteriler ve Ökaryotlar, üç-saha sisteminin (İngilizce: three domain system) temel gruplarıdır. Bakteriler gibi arkaeler de çekirdeği olmayan tek hücreli canlılardır, yani prokaryotlardır (prokaryotlar altı-âlemli sınıflandırmada Monera olarak adlandırılırlar). İlk tanımlanan arkaeler aşırı ortamlarda bulunmuş olmalarına rağmen sonradan hemen her habitatta rastlanmışlardır.
Bu üst krallığa ait tek bir organizma "arkeli" (Arkea'ye ait anlamında; İngilizce: archaean) olarak adlandırılır. Bu sözcük, sıfat olarak da kullanılır.
Tarihçe
1977'de Carl Woese ve George Fox, prokaryotları 16S rRNA dizinlerine göre sınıflandırdıkları filojenetik ağaçdaki diğer bakterilerden ayrı kümelenmelerinden dolayı arkaeleri tanımlanmışlardır. Bu iki canlı grubu başlangıçta birer âlem veya alt âlem olarak görülmüş, Arkaebakteriler ve Öbakteriler olarak adlandırılmışlardır. Woese bu grupların canlıların temel düzeyde birbirinden farklı birer kolu sayılması gerektiğini savunmuştur. Daha sonra bu kavramı daha belirginleştirmek için grupları Arkeler ve Bakteriler olarak yeniden adlandırmış ve bunların, Ökarya ile beraber canlıların üç bölgesini oluşturduğunu öne sürmüştür. (Woese'nin bu gruplara İngilizce 'bölge' anlamında domain olarak adlandırmıştır; Türkçe üst-âlem olarak da adlandırılırlar.)
Biyolojik bir terim olarak Arkea ile jeolojideki Arkean veya Arkeozoik dönemin bir ilişkisi yoktur. Arkeozoik dönem, Yer tarihinde Arke ve Bakterilerin gezegende yaşayan tek canlılar olduğu bir dönemin ismidir. Bu canlılara ait muhtemel fosiller 3,8 milyar yıl öncesine tarihlenmişlerdir.
Moleküler biyolojide temel rolü olan genetik transkripsiyon ve translasyon mekanizmaları bakterilere pek benzemeyip, çoğu bakımdan ökaryotlara benzemektedir. Örneğin arke translasyonu ökaryotik-benzeri başlatma (initiation) ve uzatma (elongasyon) faktörleri kullanır, trankripsiyonda ökaryotlardaki gibi TATA-bağlanma proteinleri ve TFIIB rol oynar. Çoğu arke tRNA ve rRNA genlerinde arkelere has intronlar bulunur ki bunlar ve ökaryotik intronlara, ne de bakteryel intronlara benz farklı kılan çeşitli başka özellikler vardır. Bakteri ve ökaryotlarda olduğu gibi arkaelerde de gliserollu fosfolipitlere sahiptirler. Ancak arke lipitlerinin üç özelliği değişiktir:
- Arke lipitlerindeki gliserolun stereokimyası bakteri ve ökaryotlardakinin tersidir. Bu, farklı bir biyosentetik yol olduğuna işarettir.
- Çoğu bakteri ve ökaryotun hücre zarları gliserol-lipit esterlerinden oluşur, oysa arkelerin zarları gliserol-lipit eterlerinden oluşur. Bakterilerde eter bağlantılı lipitler olsa dahi bunlardaki gliserol sterokimyası bakteriyel biçimdedir.
- Arke lipitleri izoprenoid birimlerden. Bu beş karbonlu bileşik bakteri ve ökaryotlardaki bazı vitaminlerde yer almasına rağmen, yalnızca arkeler onu lipitlerinin inşasında kullanırlar. Çoğunlukla bu lipitler 20 karbonlu (4 monomerden oluşmuş) veya 40 karbonlu (8 monomer) olurlar. Kırk karbonlu lipitlerin uzunluğu hücre zarının kalınlığı kadar olduğu için bazı arkelerin hücre zarında bu lipit zincirinin iki ucunda gliserol fosfat grupları bağlıdır, zar başka canlı türlerinde olduğu gibi iki lipit tabakasından değil, tek bir tabakadan oluşur. Tek tabakalı zar özellikle ısısever (termofilik) arkelerde yaygındır.
Arke hücre duvarları da bakteri ve ökaryotlarda ender görülen özelliklere sahiptir. Örneğin, çoğu arkenin hücre duvarı S-tabakası olarak adlandırılan yüzey proteinlerinden oluşur. S-tabakası bakterilerde de görülür, bazı canlılarda hücre duvarının tek bileşenidir (örneğin Planctomyces) veya peptidoglikanlı canlılarda bir dış tabaka oluşturur. Metanojenlerin bir grubu haricinde arkelerde peptidoglikan duvar yoktur. Metanojenlerde olan peptidoglikan dahi bakterilerdekinden çok farklıdır.
Arkelerin flagellası(kamçısı), bakteri flagellasına yüzeysel olarak benzese de yapı ve oluşum bakımından çok farklıdır. Bakteri flagellaları değişime uğramış bir tip III salgı sistemidir, oysa arkae flagellası tip IV pilusa benzeyip, tip II salgı sistemine benzer bir salgı sistemi kullanırlar.
Habitatları
1970'li yılların sonunda keşfedilen arkebakteriler birçok biyoloğu fazlasıyla şaşırtmıştır.Çünkü bu bakteriler aşırı sıcak, aşırı tuz gibi çok ekstrem koşullarda yaşayabilme özelliğine sahiptirler. Çoğu arke, aşırıseverdir (ekstremofil). Bazısı yüksek sıcaklıklarda, gayzerlerde veya deniz dibi sıcak su kaynaklarında oluğu gibi, çoğu zaman 100 °C'nin üstünde yaşarlar. Diğerleri çok soğuk ortamlarda, veya aşırı tuzlu, asit veya alkali ortamlarda bulunurlar. Buna karşın başka arkeler ılıman şartlarda yaşarlar (mezofil), bataklık, deniz suyu, toprak ve atık sularda bulunmuşlardır. Çoğu metanojenik bakteri geviş getiren hayvanların, insanların ve termitlerin sindirim sisteminde bulunur. Arkeler genelde diğer organizmalar için zararsızdır ve hastalık etmeni olarak bilineni yoktur.
Arkeler tercih ettikleri habitatlarına göre beş gruba ayrılırlar. Bunlar tuzsevenler (Halofiller), metanojenler, sülfür indirgeyenler, ısısevenlerdir (Termofiller) ve psikofillerdir.
- Halofiller: Aşırı tuzlu ortamlarda yaşar.En önemli özellikleri çok tuzlu alanlarda yaşayabilmeleridir. Bu tuzluluk oranı, kimi zaman ortamın tuza doyum noktasına yakın olabilir. Doğal tuz yüzeylerinde yaşarlar.Fotosentez yapabilirler.
- Metanojenler: Anaerobik ortamda yaşarlar ve metan üretirler (Oksijensiz ortamda ürerler.Enerji metabolizmalarının bir sonucu olarak metan gazı üretirler.). Arkebakterilerin büyük bir kısmını oluştururlar. Bataklıklar ve göllerin dipleri gibi oksijence fakir alanlarda, tortu tabakalarında ve hayvanların bağırsaklarında yaşarlar.Bu türler 98 °C civarında en iyi gelişimi gösterirler.84 °C’nin altındaki sıcaklıklarda ölürler. Bazı türleri ise volkanik bölgelerde sıcaklığın 110 °C olduğu sularda bulunurlar.
- Termofiller: Sıcak su kaynakları gibi yüksek sıcaklıklı yerlerde yaşarlar. Bu gruplar mutlaka moleküler genetik yöntemlerle belirlenmiş filojenilere uymayabilirler, tüm arkeleri kapsamayabilirler ve birbirlerini dışlamayabilirler. Gene de daha ayrıntılı çalışmalara başlangıç olarak faydalı sayılırlar.
- Termoasidofiller: Kemosentetik olan bu bakteriler sülfür kaynaklarında bulunmuşlardır.65-85C'lik sıcaklıkları ve pH’ın 1.0 olduğu yüksek asidik ortamda bulunurlar.
- Sülfür İndirgeyenler: Hidrojen ve genellikle volkanik kökenli inorganik sülfürü enerji kaynağı olarak kullanırlar. 85 °C sıcaklıkta yaşayabilirler.Bu bakteriler hakkında çok az şey bilinmektedir.
- Psikofiller: Bunlar aşırı soğuk ortamlarda yaşayabilen canlılardır.
Şekil
Arke hücrelerin çapları 0.1 μm ila 15 μm'nin üstü arasında değişir. Bazıları öbekleşir veya 200 μm'ye varan iplikçikler oluşturabilir. Çok çeşitli şekillere sahip olabilirler küresel, çubuk, spiral, yumrulu, yassı kare şekilli veya dikdörtgen olabilirler.
Metabolizma
Metabolizmaları çok çeşitlidir. Halobakteriler ATP üretmek için ışık kullanırlar. Ama başka gruplar gibi, elektron taşıma zinciri kullanarak fotosentez yapan bir arke yoktur. Arkelerden bulundurduğu enzimler sayesinde endüstride pek çok tepkimenin gerçekleşmesinde,atık metallerin zehirli özelliklerinin azaltılmasında,kalitesi düşük metallerin kullanılabilir hale getirilmesinde,kirlenmiş suların yeniden kullanılabilir hale getirilmesinde kullanılır.
Evrim ve sınıflandırma
Arkeler rRNA filojenetik ağaçlarına göre iki ana gruba ayrılırlar, Euryarchaeota ve Crenarchaeota. Ancak yakın yıllarda bu iki gruba ait olmayan bazı başka türler de keşfedilmiştir.
Woese, arke, bakteri ve ökaryotların ortak bir atadan (progenot) türemiş farklı evrimsel sülaleler olduğunu öne sürmüştür. Yunanca archae veya 'eski' anlamında Arke isminin seçiminin arkasında bu hipotez yatmaktadır. Daha sonra bu grupları, her biri birçok âlem içeren, bölge (domain) veya üst-âlem olarak tanımlamıştır. Bu gruplandırma sistemi çok popüler olmuş, ancak progenot fikri genel destek görmemektedir. Bazı biyologlar arkaebakteri ve ökaryotların özelleşmiş öbakterilerden türediğini öne sürmüşlerdir.
Arkea ve Ökarya arasındaki ilişki biyolojide önemli bir problem olarak sürmektedir. Yukarda belirtilen benzerlikler bir yana, birçok filogenetik ağaç bu ikisini beraber gruplandırır. Bazıları ökaryotları Crenarchaeota'lardan ziyade Euryarchaeota'lara yakın yerleştirir, hücre zarı biyokimyası aksini göstermesine rağmen. Thermatoga gibi bazı bakterilerde arke-benzeri genlerin keşfi aradaki ilişkinin tanımlanmasını zorlaştırmaktadır, çünkü yatay gen transferi olmuş olması muhtemel görünmektedir. Bazıları ökaryotların bir arkeli ile bir öbakterinin kaynaşmasıyla meydana geldiğini öne sürmüşlerdir, öyle ki birinci çekirdek, ikincisi ise sitoplazmayı oluşturmuştur. Bu hipotez genetik benzerlikleri açıklayabilmekte, ama hücre yapısını açıklamakta zorluklarla karşılaşmaktadır.
Arkelerin bakterilerden farklılıkları rRNA gen dizinlerinin karşılıştırılması sonucu ortaya çıkmıştı. Yukarıda belirtilen problemlerin bazıları, gen dizinlerine tek başına bakmak yerine artık organizmaların bütün genomlarının karşılıştırılması yoluyla çözülmeye çalışılmaktadır. 2006 Eylül ayı itibarıyla 28 arke genom dizini tamamlanmış, 28'i ise kısmen tamamlanmıştır.
Özellikleri
- Bakteriler gibi çekirdeği olmayan prokaryot, tek hücreli canlılardır.
- Bazılarında hücre duvarı bulunmaz.Bulunanlarda ise hücre duvarının kimyasal yapısı bakterilerinkinden farklıdır.
- Küresel, çubuk, spiral, dikdörtgen gibi çok çeşitli şekillere sahiptirler.
- Çok ekstrem (aşırı uç ) koşullarda yaşayabilirler.
- DNA'sı üzerinde protein kılıf bulunur (kromozom)
- Mayoz bölünme geçirmez, bölünerek çoğalır.
Arkeler üzerinde çalışmış biyologlar
- Aled Edwards, Ph.D., University of Toronto
- Carl Woese, Ph.D., University of Illinois at Urbana-Champaign
- Karl Stetter, Ph.D.,University of Regensburg, Germany
- John N. Reeve, Ph.D., Ohio State University
Kaynakça
- Howland, John L. (2000). The Surprising Archaea: Discovering Another Domain of Life. Oxford: Oxford University Press. ISBN 0-19-511183-4
- Giovannoni, S.J. and Stingl, U. (2005). Molecular diversity and ecology of microbial plankton. Nature 437: 343-348.
- Könneke, M., Bernhard, A.E., de la Torre, J.R., Walker, C.B., Waterbury, J.B. and Stahl, D.A. (2005). Isolation of an autotrophic ammonia-oxidizing marine archaeon. Nature 437: 543-546.
- Lake, J.A. (1988). Origin of the eukaryotic nucleus determined by rate-invariant analysis of rRNA sequences. Nature 331: 184–186.
- Woese, Carl R.; Fox, George E. (1977). Phylogenetic Structure of the Prokaryotic Domain: The Primary Kingdoms. Proceedings of the National Academy of Sciences of the United States of America 74 (11): 5088–5090.
- Woese, Carl R., Kandler, Otto, Wheelis, Mark L (1990). Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proceedings of the National Academy of Sciences 87 (12): 4576–4579.
Dış bağlantılar
- Archaea (İngilizce)
- ArchaeaWeb - by UNSW - Information about Archaea(İngilizce)
- Introduction to the Archaea, ecology, systematics and morphology29 Ekim 2004 tarihinde Wayback Machine sitesinde arşivlendi.(İngilizce)
- Archaea at The Encyclopedia of Astrobiology, Astronomy, & Spaceflight(İngilizce)
- Extremophiles Bioprospecting for antimicrobials, Dr Sarah Maloney23 Nisan 2006 tarihinde Wayback Machine sitesinde arşivlendi. (İngilizce)
- BBC News July 21, 1999: Toughest bug reveals genetic secrets5 Şubat 2007 tarihinde Wayback Machine sitesinde arşivlendi. Citat: "...It [Pyrococcus abyssi] likes conditions that the vast majority of other organisms would find impossible to live in. It thrives best at temperatures of about 103 degrees [Celsius] and under pressures of about 200 atmospheres...."(İngilizce)
- Pyrococcus abyssi Home page at Genoscope24 Ekim 2006 tarihinde Wayback Machine sitesinde arşivlendi.(İngilizce)
- Browse any completed archaeal genome at UCSC5 Ocak 2005 tarihinde Wayback Machine sitesinde arşivlendi.(İngilizce)
- 3D structures of proteins from archaebacterial membranes8 Eylül 2006 tarihinde Wayback Machine sitesinde arşivlendi. (İngilizce)
