DNA polimeraz
DNA polimeraz, DNA ikileşmesini sağlayan bir enzim. Bu enzimler bir DNA ipliğini kalıp olarak kullanır, onu okuyup, onun boyunca deoksiribonükleotitlerin polimerizasyonunu katalizler. Yeni polimerleşmiş molekül kalıp ipliği tamamlayıcıdır, ve kalıp ipliğin eski eşi ile aynı yapıya sahiptir.
| DNA-yönlendirmeli DNA polimeraz | |||||||
|---|---|---|---|---|---|---|---|
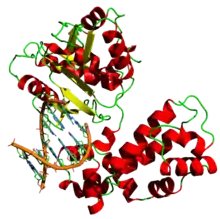 | |||||||
| İnsan DNA polimeraz beta'nın DNA'ya bağlanan motifleri (helix-turn-helix ) (7ICG pdb dosyasından oluşturulmuştur) | |||||||
| Tanımlayıcılar | |||||||
| EC numarası | 2.7.7.7 | ||||||
| CAS numarası | 9012-90-2 | ||||||
| Veritabanları | |||||||
| IntEnz | IntEnz view | ||||||
| BRENDA | BRENDA entry | ||||||
| ExPASy | NiceZyme view | ||||||
| KEGG | KEGG entry | ||||||
| MetaCyc | metabolic pathway | ||||||
| PRIAM | profile | ||||||
| PDB structures | RCSB PDB PDBe PDBsum | ||||||
| Gene Ontology | AmiGO / EGO | ||||||
| |||||||
DNA polimeraz bir holoenzim olarak sayılır çünkü doğru işlev verebilmek için bir magnezyum iyonuna gerek duyar. Magnezyum iyonunun yokluğunda ona apoenzim olarak değinilir.
DNA polimeraz tek iplikli bir DNA'ya bağlanarak DNA ikileşmesini başlatır. RNA polimerazdan farklı olarak, DNA polimeraz sentezlediği yeni ipliği sadece nükleotitlerden başlayarak uzatamaz, ancak mevcut bir DNA zincirini uzatabilir. Bu yüzden zincir sentezinin başı için yardımcı enzimlere gerek duyar. DNA polimerazların bazı türleri hatalı ekledikleri nükleotitleri fark edip onları tamir etmelerini sağlayan bir eksonükleaz yeteneğine sahiptir.
İşlev
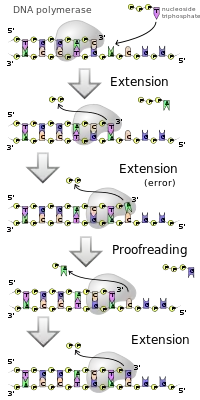
DNA polimeraz yeni uzayan bir ipliğin sadece 3' ucuna serbest nükleotitler bağlayabilir. Bunun sonucu olarak yeni ipliğin uzaması 5'->3' yönünde olur. Bilinen DNA polimerazlar arasında yeni bir zinciri tamamen başlangıçtan oluşturma yeteneği olan yoktur. DNA polimeraz sadece mevcut bir 3'-OH grubuna bir nükleotit ekleyebilir, ve dolayısıyla ilk nükleotidi ekleyebileceği bir başlatıcıya ("primer"e) gerek duyar. Primerler RNA ve DNA bazlarından oluşur, ilk iki baz RNA'dır, ve primaz adlı başka bir enzim tarafından üretilirler. Helikaz denen bir enzim ise, DNA'nın ikili sarmalını ters yönde burarak gevşetir ve onu tek iplikli bir şekil almasını sağlar. Böylece DNA, yarı saklı ikileşmeye hazır hale gelir.
Hata tamiri bazı polimerazların bir özelliği olmakla beraber hepsinde görülmez. Bu süreç, yeni sentezlenen DNA'daki hataları düzeltir. Hatalı bir baz çifti fark edilince, DNA polimeraz yönünü değiştirip bir baz çifti geriye gider. Enzimin 3'->5' eksonükleaz aktivitesi hatalı bazları kesilip çıkartılmasını sağlar. Bazın çıkartılmasından sonra polimeraz doğru bazı uzayan zincire dahil eder ve ikileşme devam eder.
Türler arasında çeşitlilik
DNA polimerazların yapıları evrim zarfında korunmuştur, yani bir türden başka bir türe bu enzimler arasında pek az bir fark vardır. Korunmuş yapılar, hücre için önemli, değiştirilemez işlevlere işaret eder, dolayısıyla bunların devamının sürdürülmesi evrimsel bir avantaj sağlar.
Bazı virüsler ayrıca viral DNA'yı çeşitli mekanizmalarla ikileştirebilen özel DNA polimerazlar kodlar. Retrovirüsler ters transkriptaz adı verilen olağandışı bir polimeraz kodlar, bu RNA-bağımlı bir DNA polimerazdır. Bunlar DNA'yı polimerleştirmek için bir RNA kalıp kullanılır.
DNA polimeraz aileleri
DNA dizi benzerliklerine dayanarak DNA polimerazlar yedi farklı aileye bölünmüşlerdir: A, B, C, D, X, Y, ve RT.
A ailesi
A ailesi polimerazlar hem ikileşme hem de tamir polimerazları içerir. Bu aileye ait ikileştirici üyeleri arasında çok çalışılmış olan T7 DNA polimeraz ve ayrıca ökaryotik mitokondrial DNA Polimeraz γ bulunur. Tamir polimerazları arasında E. coli DNA pol I, Thermus aquaticus pol I, ve Bacillus stearothermophilus pol I bulunur. Bu tamir polimerazları kesip çıkarma (eksizyon) tamiri ve ikileşme çatalında gecikmeli iplik (İng. lagging strand) sentezi sırasında Okazaki parçalarının işlenmesi ile ilgilidir.
B ailesi
B ailesi polimerazlar başlıca ikileşmeli polimerazlar içerir; aralarında ökaryotik polimerazların baçlıcaları olan α (alfa), δ (delta), ε (epsilon) ve ayrıca DNA polimeraz ζ (zeta) bulunur. B familyası ayrıca bazı bakteri ve bakteriyofajlar tarafından kodlanmış DNA polimerazlar da içerir, bunların arasında en iyi tanımlanmış olanlar T4, Phi29, ve RB69 bakteriyofajlarına ait olanlardır. Bu enzimler hem öncü hem de gecikmeli zincir senteziyle ilişkilidirler. B ailesi DNA polimerazların en tanımlayıcı özelliği, ikileşme sırasındaki yüksek hatasızlık oranıdır. Bunların çoğunun güçlü bir 3'-5' eksonükleaz aktivitesi vardır (DNA polimeraz α ve ζ hariç, bunların prova okuma aktivitesi yoktur).
C ailesi
C ailesi polimerazlar en başlıca bakteri kromozom ikileştirme enzimleridir. E. coli 'nin DNA Polimeraz III alfa altbiriminin bilinen herhangi bir eksonükleaz aktivitesi yoktur. Bir diğer altbirim, epsilon, kromozom ikileşmesi için kullanılan 3'->5' eksonükleaz aktivitesine sahiptir.
D ailesi
D ailesi polimerazlar halen iyi tanımlanmış değllerdir. Tüm bilinen örnekleri Arkelerin Euryarchaeota altbölümünde bulunur, bunların ikileşme polimerazı olduğu düşünülmektedir.
X ailesi
X ailesi iyi bilinen ökaryotik polimeraz pol β (beta), ayrıca ökaryotik polimerazlar pol σ (sigma), pol λ (lambda), pol μ (mu), ve terminal deoksinükleotidil transferaz (TdT) içerir. Pol β, baz eksiyon tamiri için gereklidir, bu bazsız DNA bölgelerinin tamiri için gerekli bir tamir yoludur. Pol λ ve Pol μ, homolog olmayan uç birleştirme ile ilişkilidir, bu, çift zincirli DNA kırıklarını birleştirmek için bir mekanizmadır. TdT sadece lenf dokularında bulunur, immün çeşitlilik oluşturmaya yarayan V(D)J rekombinasyonu sırasında meydana gelen çift zincirli DNA kırıklarına nükleotitler ekler. Saccharomyces cerevisiae mayasında tek bir Pol X polimeraz bulunur, Pol4, bu homolog olmayan DNA uç birleştirmesinde görev alır.
Y ailesi
Y ailesi polimerazları, zarar görmemiş kalıpları kopyalarken hata oranlarının yüksek olması, ama zarar görmüş DNA'yı ikileştirebilme özellikleriyle diğer polimerazlardan farklılık gösterirler. Bu ailenin üyeleri dolayısıyla lezyon-aşırı sentez (İng. "translesion synthesis"; TLS) polimerazları olarak adlandırılırlar. Lezyona bağlı olarak zarar görmüş bölgeyi ya hatasız veya hataya eğilimli şekilde geçebilirler. Kseroderma pigmentosum hastalığının XPV çeşidi hastalarda Pol η (eta) kodlayan gende mutasyonlar bulunur, bu polimeraz normalde morötesi lezyonlarda hatasız çalışır. XPV hastalarında, bu polimeraz yerine hataya eğilimli alternatif polimerazlar, örneğin Polζ (zeta) (polimeraz ζ bir B ailesi polimerazıdır) çalışır, bunlar da bu hastalarda kansere yol açan hatalara neden olur. İnsanlarda bu ailenin diğer üyeleri Pol ι (iota), Pol κ (kappa), ve Rev1 (terminal deoksisitidil transferaz)'dir. E.coli 'de iki TLS polimeraz, Pol IV (DINB) ve PolV (UmuD'2C), vardır.
RT ailesi
Ters transkriptaz ailesi hem retrovirüslerden hem de ökaryotik polimerazlardan örnekler içerir. Ökaryotik polimerazlar telomerazlardan ibarettir. Bu polimerazlar bir RNA kalıptan DNA zinciri sentezlerler.
Canlı türlerine göre gruplandırma
Prokaryotik DNA polimerazlar
Bakterilerde 5 DNA polimerazın varlığı bilinmektedir:
- Pol I: DNA tamirinde görev alır. Hem 5'->3' polimeraz aktivitesi hem de 5'->3' eksonükleaz activitesi vardır (RNA primerleri çıkarmak için). DNA'ya bağlanması ile ondan ayrışması arasında ortalama 20 nükleotit ekler, yani ilerleyiciliği (İng. processivity) azdır. ayrıca 3'->5' eksonükleaz aktivitesi vardır.
- Pol II: Zarar görmüş DNA'nın ikileşmesinde görev alır; 3'->5' eksonükleaz aktivitesi vardır.
- Pol III: Bakterilerin esas polimerazıdır (DNA ikileşmesinde DNA'yı uzatır); 3'->5' eksonükleaz prova okuma yeteneği vardır.
- Pol IV: Y ailesi üyesi bir DNA polimerazdır
- Pol V: Y ailesi uyesidir; DNA'nın zarar görmüş bölgelerini aşmaya yarar.
Arke DNA polimerazları
Arkelerde iki tip DNA polimeraz vardır.
- Polimeraz B: B tipi polimerazların hepsinin 3'->5' polimeraz ve 5'->3' eksonükleaz aktivitesi vardır. Tek alt birimden oluşurlar.
- Polimeraz D: Euryarchaeota'larda görülen bu DNA polimerazın iki alt birimi vardır. Büyük altbirim DP2 polimeraz aktivitesine sahiptir ama bunun çalışabilmesi için küçük DP1 biriminin de bulunmsı gerekir.
Ökaryotik DNA polimerazlar
Ökaryotların en az 15 DNA polimerazı vardır:[1]
- Pol α (eşanlamlı adları DNA primaz, RNA polimeraz): primaz olarak etkir (bir RNA primer sentezler), ve sonra bu primeri DNA nükleotitleri ile uzatır. Yaklaşık 20 nükleotit sonra uzama reaksiyonu Pol δ (gecikmeli iplikte) veya ε (öncü iplikte) uzatma işlemini devralır.[2]
- Pol β: DNA tamirinde görev alır, baz eksizyon tamiri ve boşluk doldurması yapar.
- Pol γ: Mitokondrial DNA'yı ikileştirir ve tamir eder; 3'->5' eksonükleaz aktivitesiyle prova okuması yapar.
- Pol δ: Yüksek derecede ilerleyicidir ve 3'->5' eksonükleaz aktivitesiyle prova okuması yapar. İkileşme çatalındaki gecikmeli iplik sentezinden sorumlu esas polimeraz olduğu düşünülmektedir ama bu konu hâlâ tartışmalıdır.[3]
- Pol ε: Bu da çok ilerleyici bir enzimdir ve 3'->5' eksonükleaz aktivitesine sahiptir. Pol δ ile yakın akrabalığı vardır, ve öncü iplik (İng. leading strand) sentezinden sorumlu esas polimeraz olduğu düşünülmektedir[4] ancak bunun rolü hakkında da tartışma sürmektedir[3].
- η, ι, κ, ve Rev1, Y ailesi DNA polimerazlarıdır. Bu polimerazlar DNA zarar bölgelerinin atlanmasında işlev görürler.[5]
- Başka ökaryotik polimerazlar da bilinmektedir ama bunlar iyi tanımlanmamıştır: θ, λ, φ, σ, ve μ. Başka polimerazlar daha vardır ama bunlar isimlendirilmesi halen yerleşmemiştir.
Bu polimerazların hiçbiri primerleri çıkaramaz (5'->3' eksonükleaz aktivitesi yoktur); bu fonksiyon başka enzimler tarafından sağlanır. Sadece uzamadan sorumlu polimerazların (γ, δ ve ε) prova okuma yeteneği (3'->5' eksonükleaz aktivitesi) vardır.
Biyoteknolojide önemi
Laboratuvarda DNA polimerazlar çoğu zaman polimeraz zincir reaksiyonu için kullanılırlar. Bu amaç için kullanılan polimerazların yüksek ısıya dayanıklı olması ve hata düzeltme (prova okuma) fonksiyonu olması gereklidir, bu yüzden yüksek ısıda yaşayabilen canlıların (örneğin Thermus aquaticus) DNA polimerazı veya onun yapay türevleri kullanılır. DNA polimeraz ayrıca DNA dizilemesinde de kullanılır.
Ayrıca bakınız
Kaynakça
- I. Hubscher, U.; Maga, G.; Spadari, S. (2002). "Eukaryotic DNA polymerases". Annual Review of Biochemistry. Cilt 71. ss. 133-63. doi:10.1146/annurev.biochem.71.090501.150041. PMID 12045093.
- J. M. Berg; J. L. Tymoczko; L. Stryer "Biochemie", Springer, Heidelberg/Berlin 2003
- Scott D McCulloch; Thomas A Kunkel (Ocak 2008). "The fidelity of DNA synthesis by eukaryotic replicative and translesion synthesis polymerases". Cell Research. Cilt 18. ss. 148-161. doi:10.1038/cr.2008.4. PMID 18166979.
- Pursell, Z.F.; ve diğerleri. (2007). "Yeast DNA Polymerase ε Participates in Leading-Strand DNA Replication". Science. Cilt 317. ss. 127-130. doi:10.1126/science.1144067. PMID 17615360.
- I. Prakash, S.; Johnson, R. E.; Prakash, L. (2005). "Eukaryotic translesion synthesis DNA polymerases: specificity of structure and function". Annual Review of Biochemistry. Cilt 74. ss. 317-53. doi:10.1146/annurev.biochem.74.082803.133250. PMID 15952890.